

Endoglin regulates PI3-kinase/Akt trafficking and signaling to alter endothelial capillary stability during angiogenesis
- PMID: 22593212
- PMCID: PMC3386206
- DOI: 10.1091/mbc.E11-12-0993
Endoglin regulates PI3-kinase/Akt trafficking and signaling to alter endothelial capillary stability during angiogenesis
Abstract
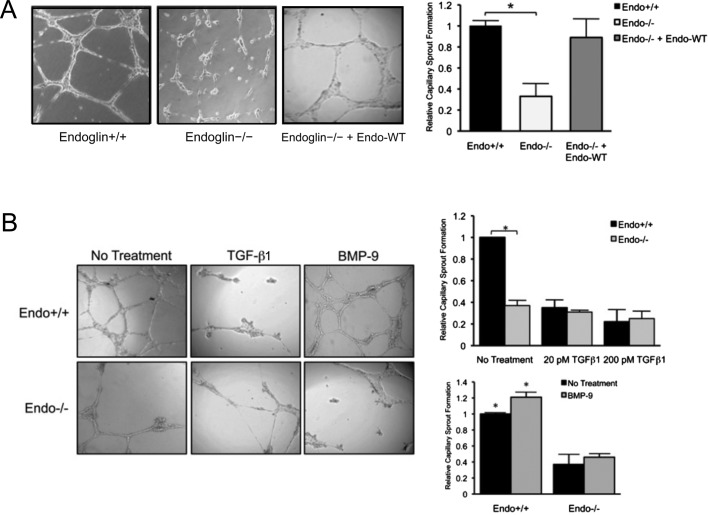
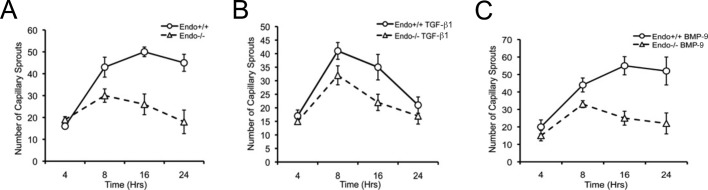
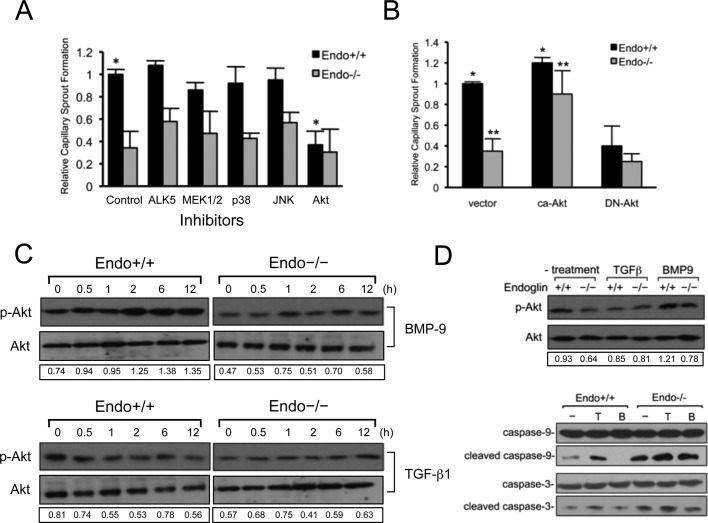
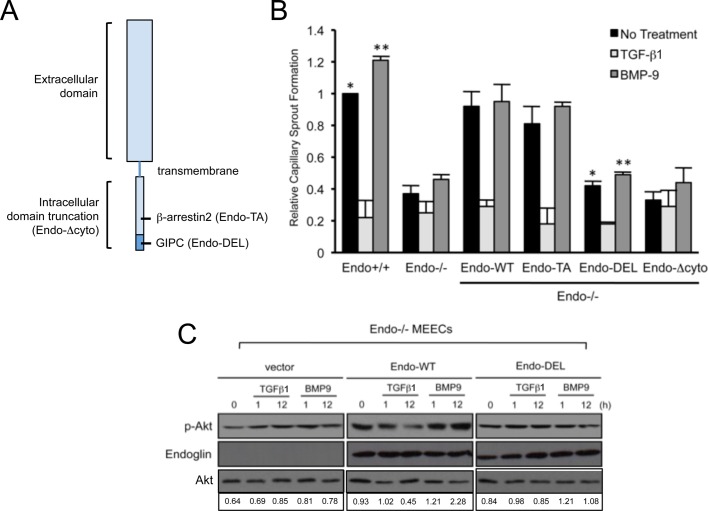
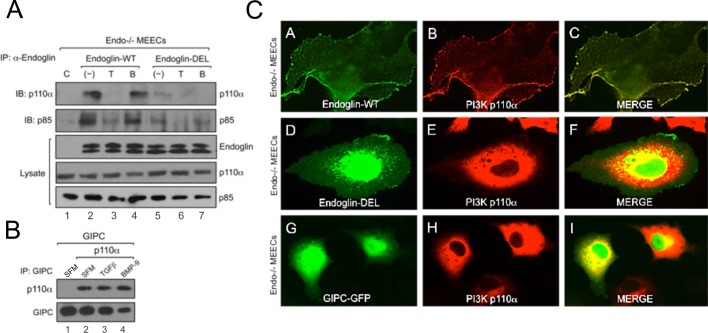
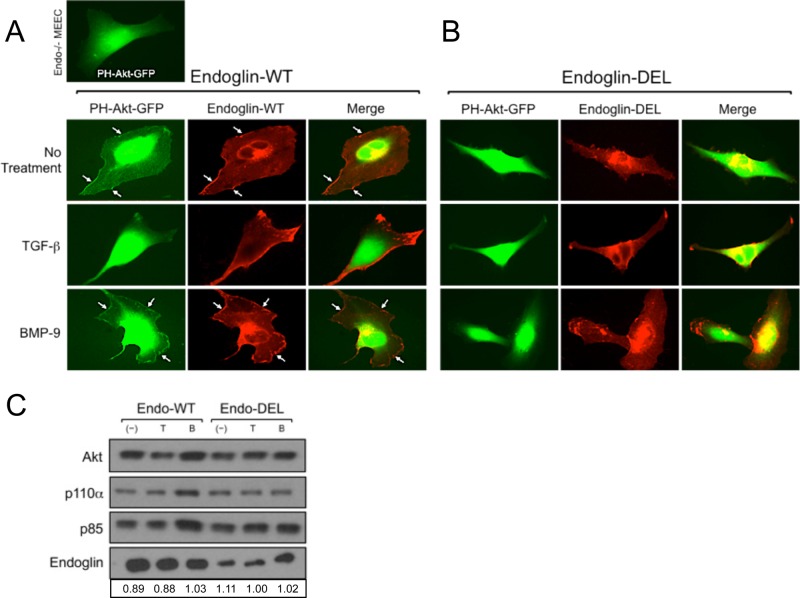
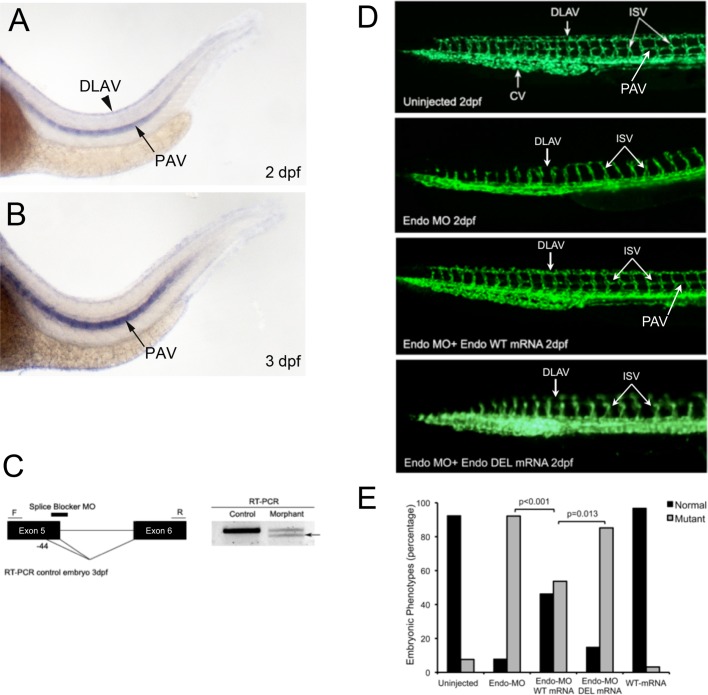
Endoglin (CD105) is an endothelial-specific transforming growth factor β (TGF-β) coreceptor essential for angiogenesis and vascular homeostasis. Although endoglin dysfunction contributes to numerous vascular conditions, the mechanism of endoglin action remains poorly understood. Here we report a novel mechanism in which endoglin and Gα-interacting protein C-terminus-interacting protein (GIPC)-mediated trafficking of phosphatidylinositol 3-kinase (PI3K) regulates endothelial signaling and function. We demonstrate that endoglin interacts with the PI3K subunits p110α and p85 via GIPC to recruit and activate PI3K and Akt at the cell membrane. Opposing ligand-induced effects are observed in which TGF-β1 attenuates, whereas bone morphogenetic protein-9 enhances, endoglin/GIPC-mediated membrane scaffolding of PI3K and Akt to alter endothelial capillary tube stability in vitro. Moreover, we employ the first transgenic zebrafish model for endoglin to demonstrate that GIPC is a critical component of endoglin function during developmental angiogenesis in vivo. These studies define a novel non-Smad function for endoglin and GIPC in regulating endothelial cell function during angiogenesis.
Figures
References
-
- Cheifetz S, Bellon T, Cales C, Vera SC, Bernabeu C, Massague J, Letarte M. Endoglin is a component of the transforming growth factor-beta receptor system in human endothelial cells. J Biol Chem. 1992;267:19027–19030. - PubMed
-
- Conley B, Koleva R, Simth J, Kacer D, Zhang D, Bernabeu C, Vary CP. Endoglin controls cell migration and composition of focal adhesions: function of the cytosolic domain. J Biol Chem. 2004;279:27440–27449. - PubMed
-
- David L, Feige JJ, Bailly S. Emerging role of bone morphogenetic proteins in angiogenesis. Cytokine Growth Factor Rev. 2009;20:203–212. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
