

Kif18A and chromokinesins confine centromere movements via microtubule growth suppression and spatial control of kinetochore tension
- PMID: 22595673
- PMCID: PMC3356572
- DOI: 10.1016/j.devcel.2012.02.013
Kif18A and chromokinesins confine centromere movements via microtubule growth suppression and spatial control of kinetochore tension
Abstract
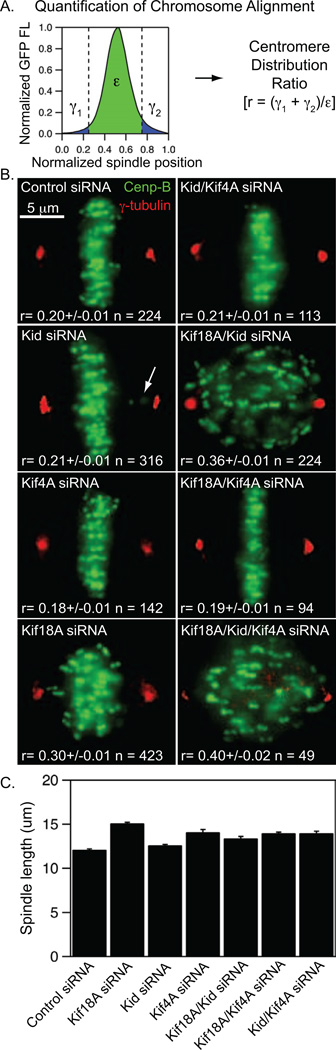
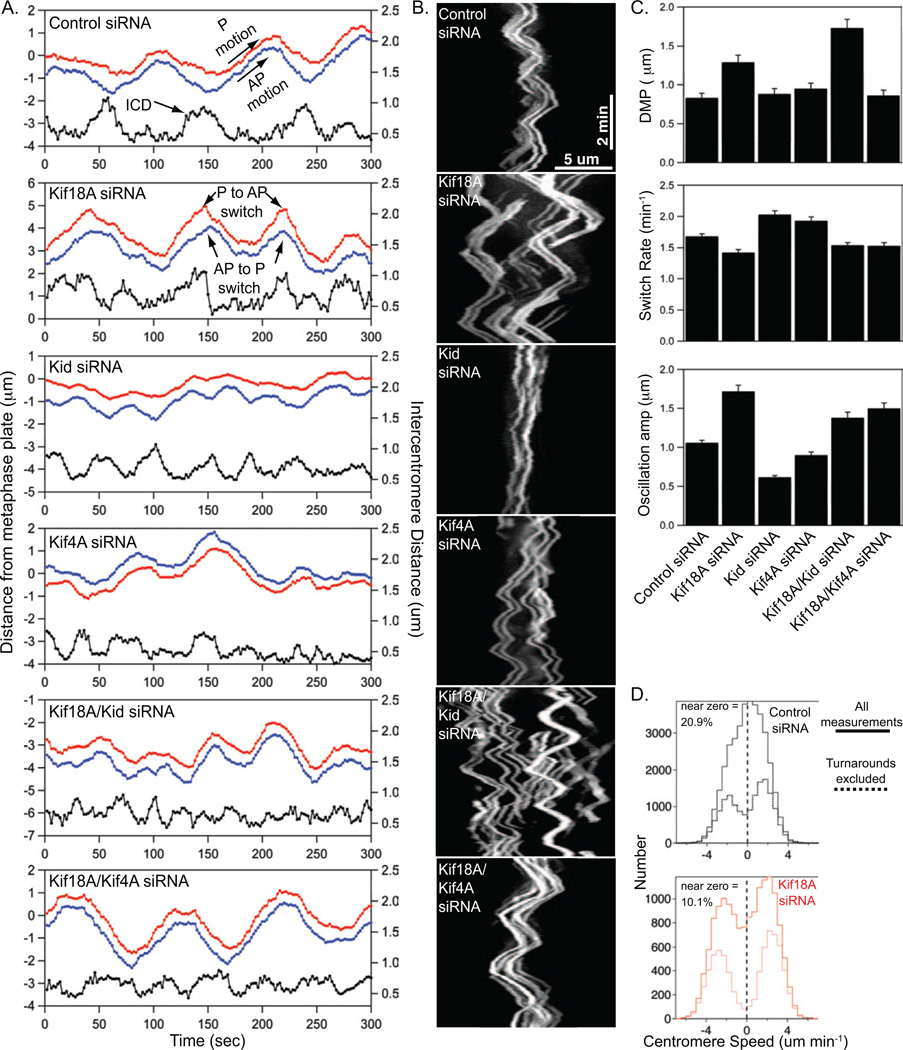
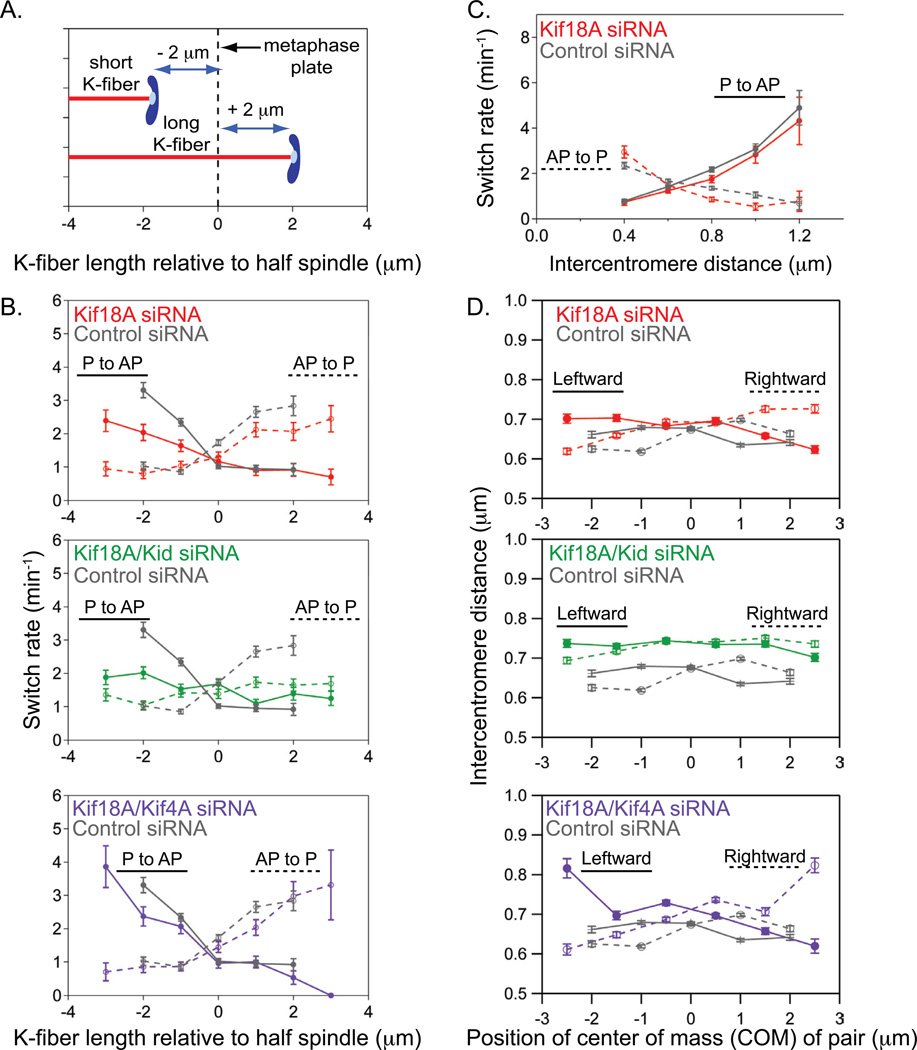
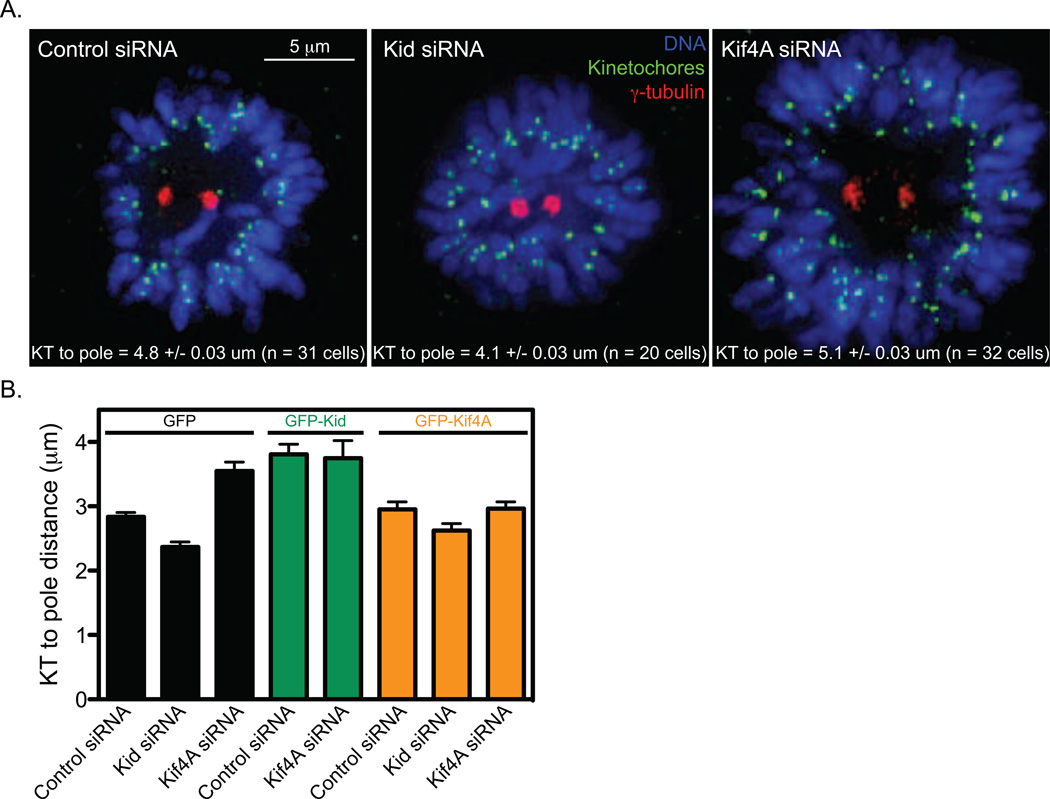
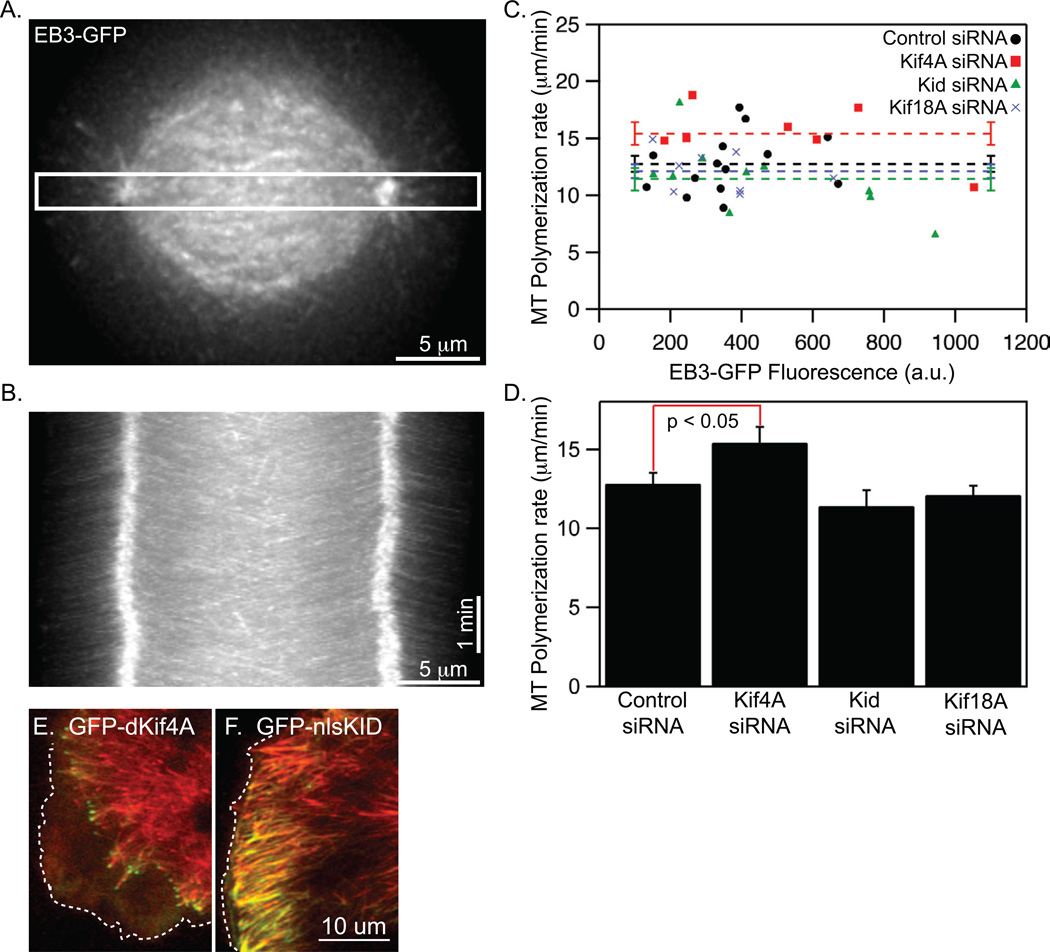
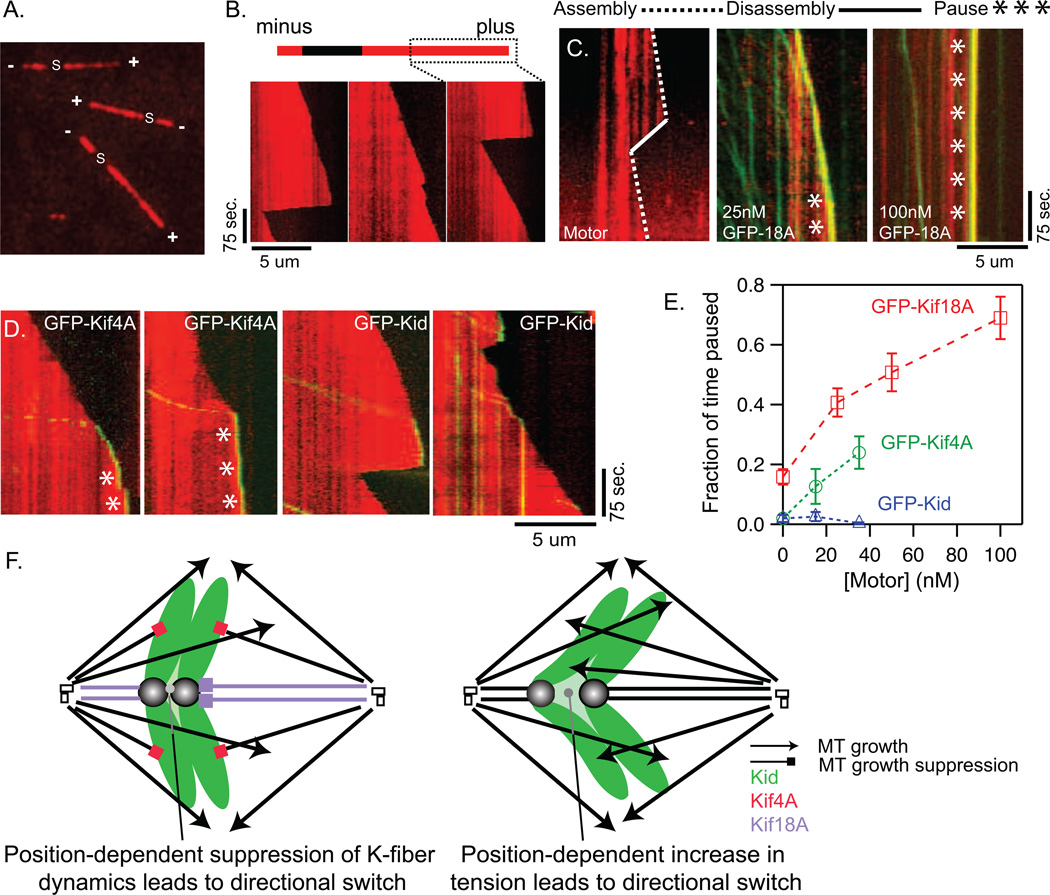
Alignment of chromosomes at the metaphase plate is a signature of cell division in metazoan cells, yet the mechanisms controlling this process remain ambiguous. Here we use a combination of quantitative live-cell imaging and reconstituted dynamic microtubule assays to investigate the molecular control of mitotic centromere movements. We establish that Kif18A (kinesin-8) attenuates centromere movement by directly promoting microtubule pausing in a concentration-dependent manner. This activity provides the dominant mechanism for restricting centromere movement to the spindle midzone. Furthermore, polar ejection forces spatially confine chromosomes via position-dependent regulation of kinetochore tension and centromere switch rates. We demonstrate that polar ejection forces are antagonistically modulated by chromokinesins. These pushing forces depend on Kid (kinesin-10) activity and are antagonized by Kif4A (kinesin-4), which functions to directly suppress microtubule growth. These data support a model in which Kif18A and polar ejection forces synergistically promote centromere alignment via spatial control of kinetochore-microtubule dynamics.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Antonio C, Ferby I, Wilhelm H, Jones M, Karsenti E, Nebreda AR, Vernos I. Xkid, a chromokinesin required for chromosome alignment on the metaphase plate. Cell. 2000;102:425–435. - PubMed
-
- Bieling P, Kronja I, Surrey T. Microtubule Motility on Reconstituted Meiotic Chromatin. Curr Biol. 2010a - PubMed
-
- Bieling P, Telley IA, Surrey T. A Minimal Midzone Protein Module Controls Formation and Length of Antiparallel Microtubule Overlaps. Cell. 2010b;142:420–432. - PubMed
-
- Bringmann H, Skiniotis G, Spilker A, Kandels-Lewis S, Vernos I, Surrey T. A kinesin-like motor inhibits microtubule dynamic instability. Science. 2004;303:1519–1522. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
