

The origin of extracellular fields and currents--EEG, ECoG, LFP and spikes
- PMID: 22595786
- PMCID: PMC4907333
- DOI: 10.1038/nrn3241
The origin of extracellular fields and currents--EEG, ECoG, LFP and spikes
Abstract
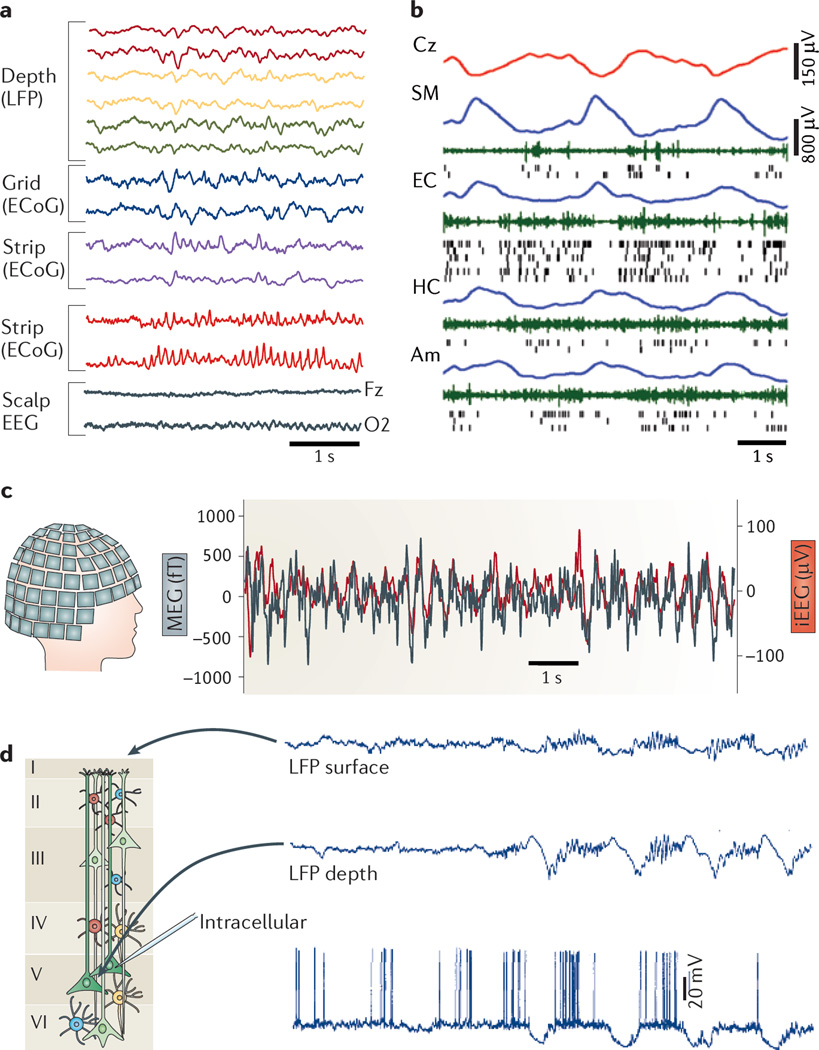
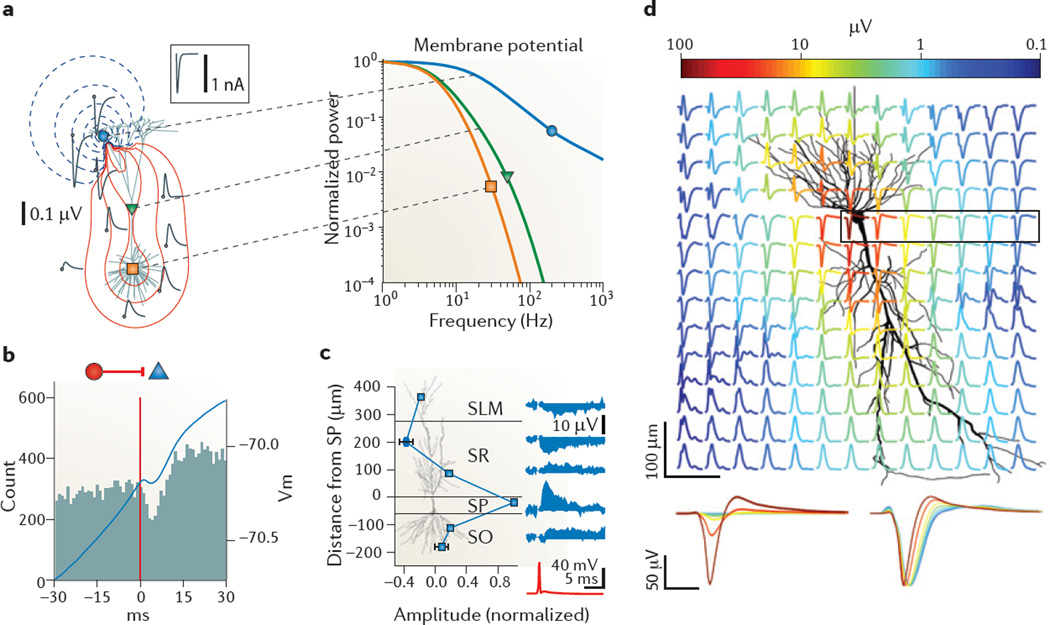
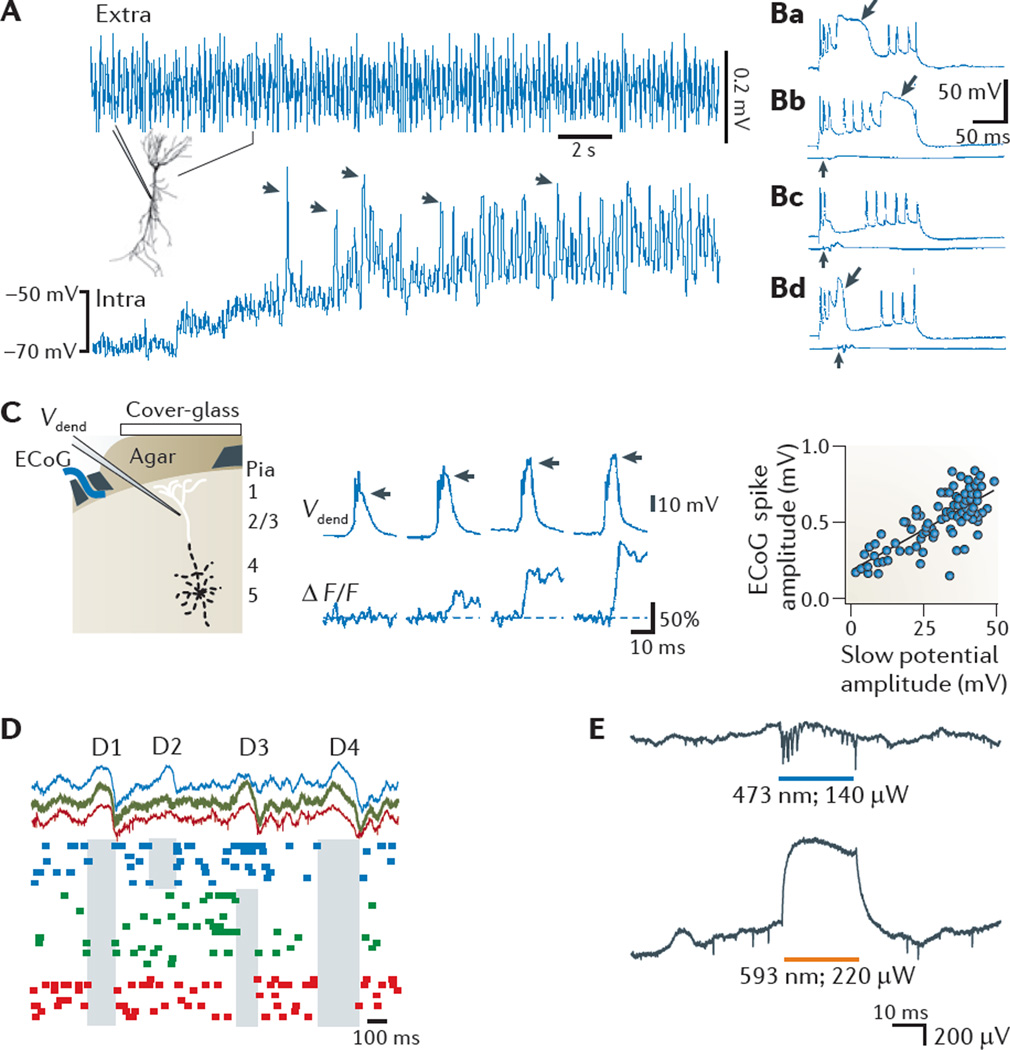
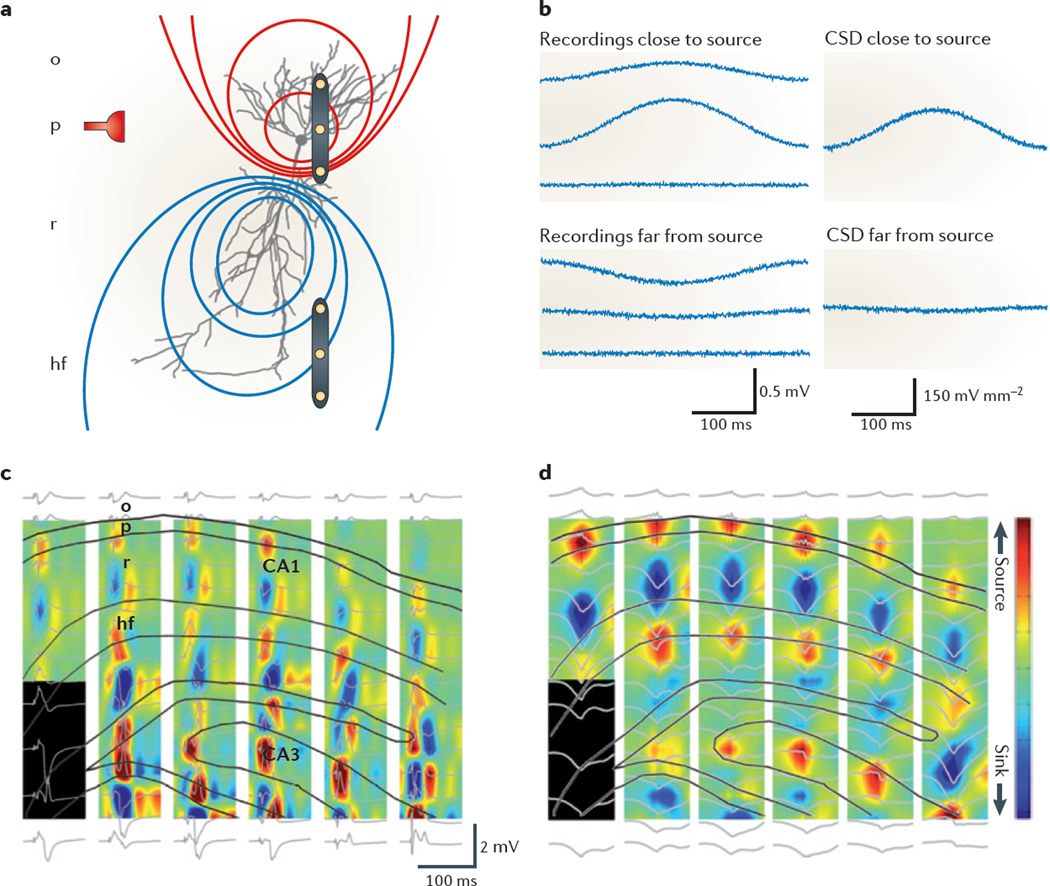
Neuronal activity in the brain gives rise to transmembrane currents that can be measured in the extracellular medium. Although the major contributor of the extracellular signal is the synaptic transmembrane current, other sources--including Na(+) and Ca(2+) spikes, ionic fluxes through voltage- and ligand-gated channels, and intrinsic membrane oscillations--can substantially shape the extracellular field. High-density recordings of field activity in animals and subdural grid recordings in humans, combined with recently developed data processing tools and computational modelling, can provide insight into the cooperative behaviour of neurons, their average synaptic input and their spiking output, and can increase our understanding of how these processes contribute to the extracellular signal.
Figures
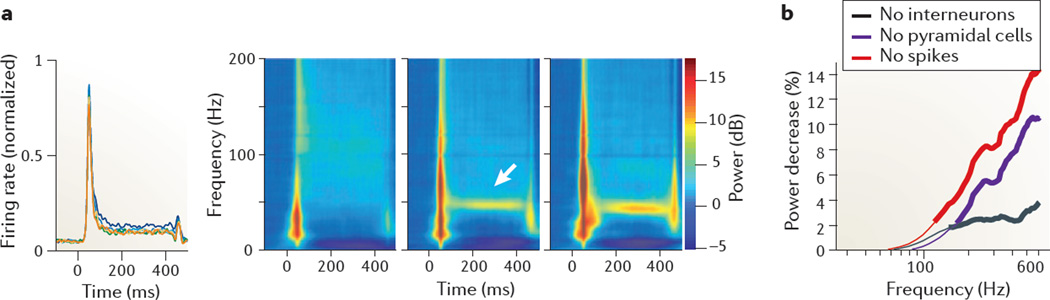
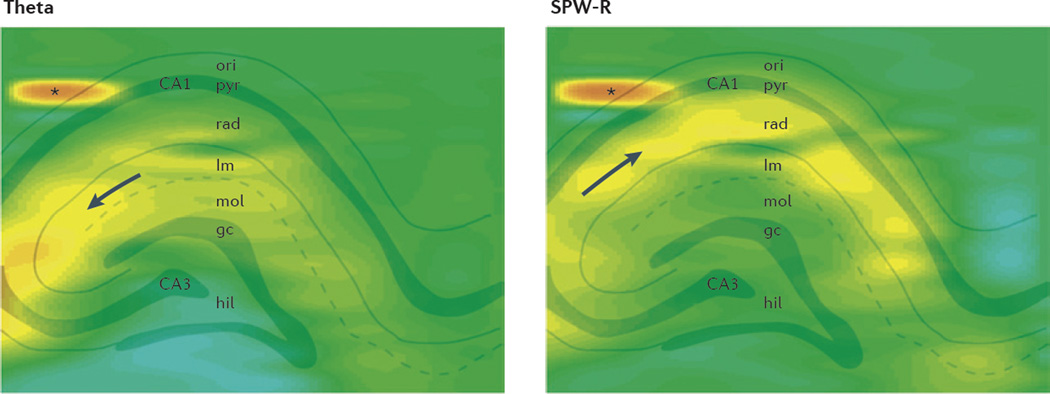
References
-
-
Petsche H, Pockberger H, Rappelsberger P. On the search for the sources of the electroencephalogram. Neuroscience. 1984;11:1–27. An excellent review of the sources of the local field.
-
-
-
Hämäläinen M, Hari R, Ilmoniemi RJ, Knuutila J, Lounasmaa OV. Magnetoencephalography — theory, instrumentation, and applications to noninvasive studies of the working human brain. Rev. Mod. Phys. 1993;65:413–497. An exhaustive review of and excellent tutorial on the theory and methods of MEG.
-
-
- Buzsáki G, Draguhn A. Neuronal oscillations in cortical networks. Science. 2004;304:1926–1929. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
