

Multiple spectral inputs improve motion discrimination in the Drosophila visual system
- PMID: 22605779
- PMCID: PMC6528803
- DOI: 10.1126/science.1215317
Multiple spectral inputs improve motion discrimination in the Drosophila visual system
Abstract
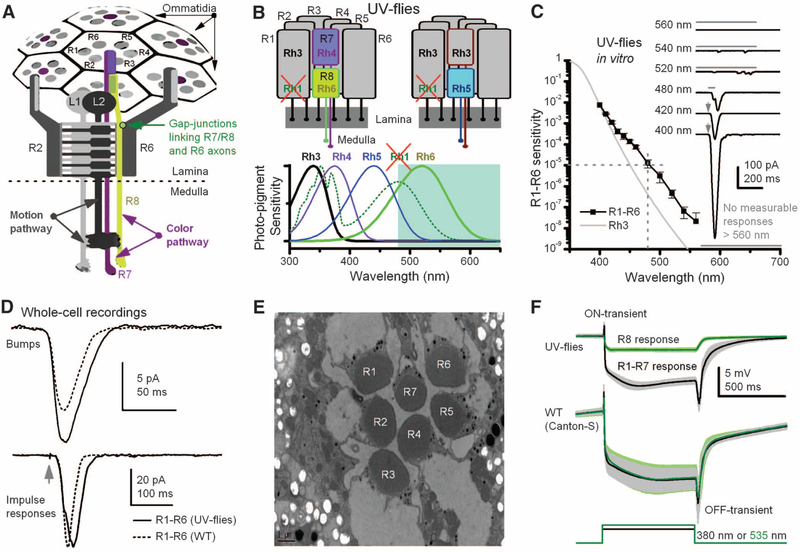
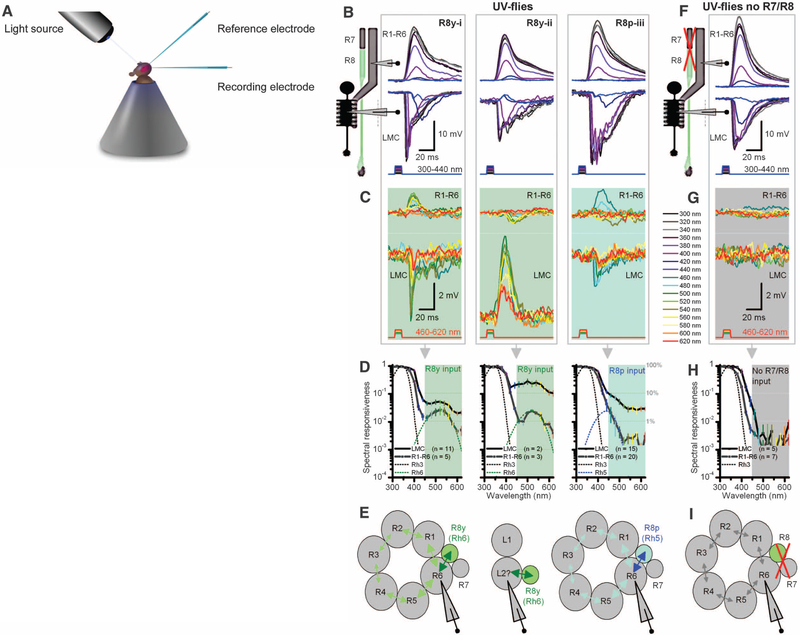
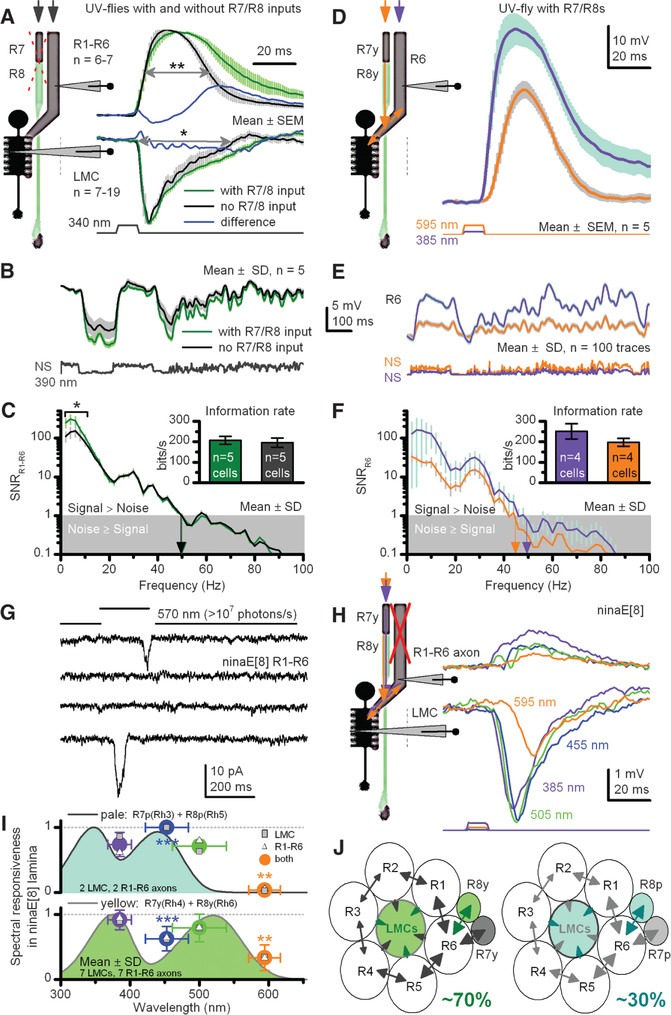
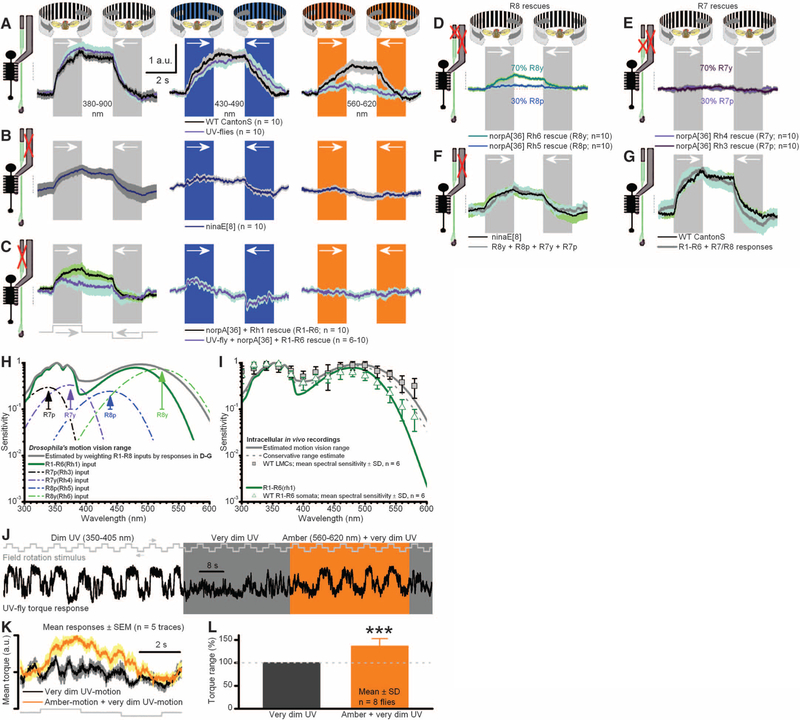
Color and motion information are thought to be channeled through separate neural pathways, but it remains unclear whether and how these pathways interact to improve motion perception. In insects, such as Drosophila, it has long been believed that motion information is fed exclusively by one spectral class of photoreceptor, so-called R1 to R6 cells; whereas R7 and R8 photoreceptors, which exist in multiple spectral classes, subserve color vision. Here, we report that R7 and R8 also contribute to the motion pathway. By using electrophysiological, optical, and behavioral assays, we found that R7/R8 information converge with and shape the motion pathway output, explaining flies' broadly tuned optomotor behavior by its composite responses. Our results demonstrate that inputs from photoreceptors of different spectral sensitivities improve motion discrimination, increasing robustness of perception.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- Z01 HD008776/ImNIH/Intramural NIH HHS/United States
- BB/H013849/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/D001900/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- ZIA HD008776/ImNIH/Intramural NIH HHS/United States
- BB/G006865/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
