

Bmi1 confers resistance to oxidative stress on hematopoietic stem cells
- PMID: 22606246
- PMCID: PMC3350495
- DOI: 10.1371/journal.pone.0036209
Bmi1 confers resistance to oxidative stress on hematopoietic stem cells
Abstract
Background: The polycomb-group (PcG) proteins function as general regulators of stem cells. We previously reported that retrovirus-mediated overexpression of Bmi1, a gene encoding a core component of polycomb repressive complex (PRC) 1, maintained self-renewing hematopoietic stem cells (HSCs) during long-term culture. However, the effects of overexpression of Bmi1 on HSCs in vivo remained to be precisely addressed.
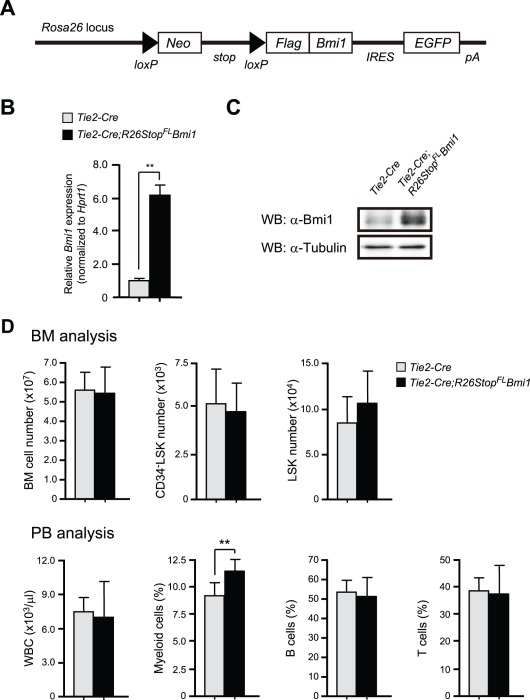
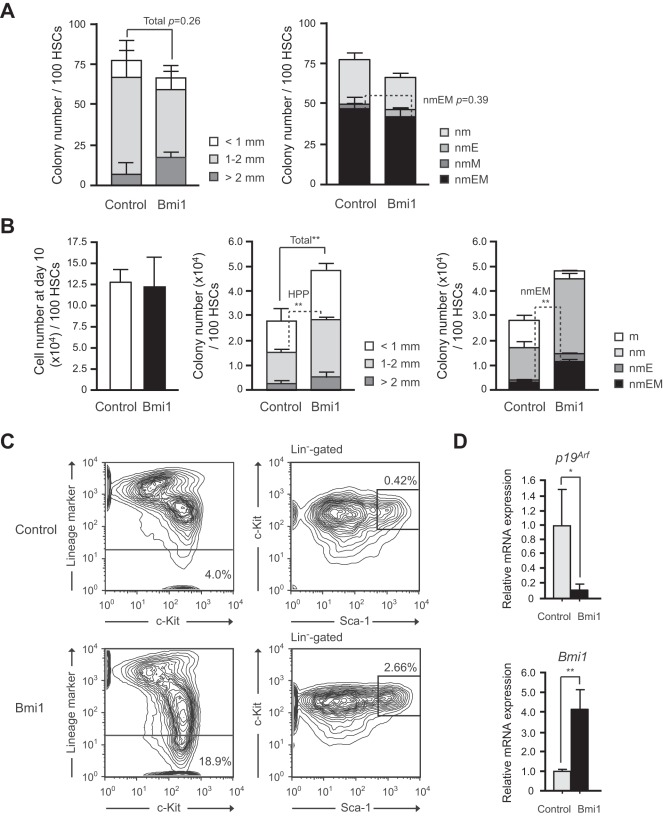
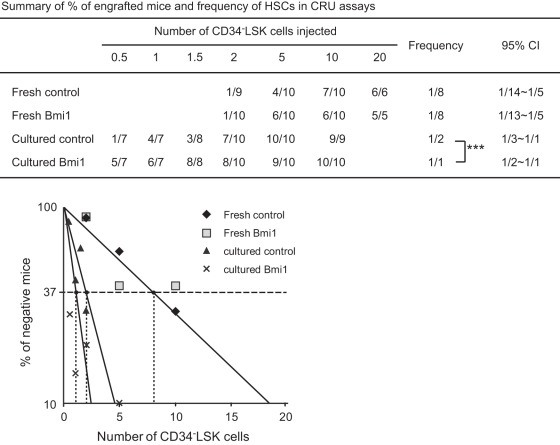
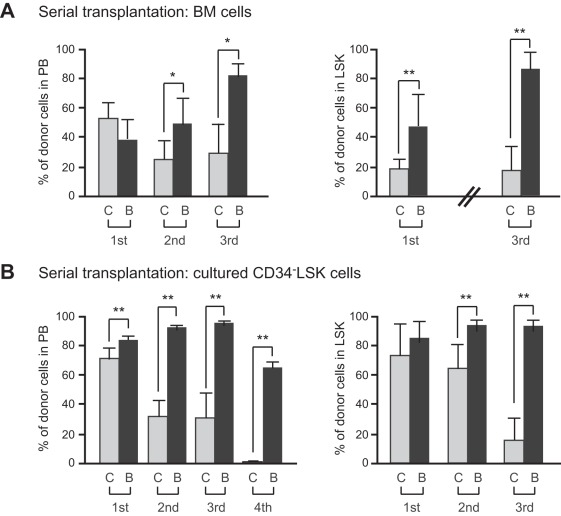
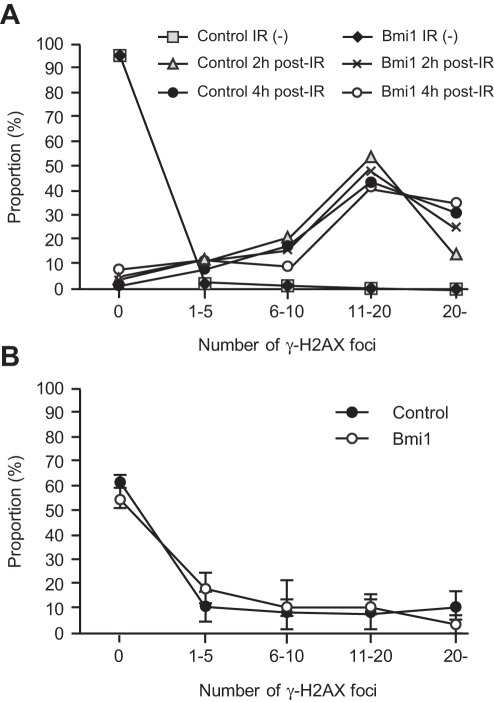
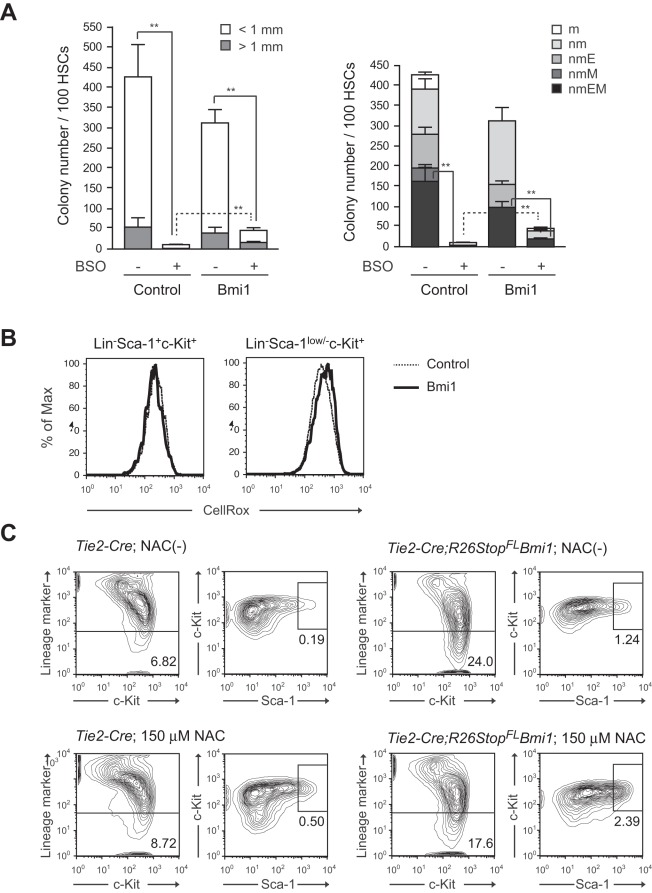
Methodology/principal findings: In this study, we generated a mouse line where Bmi1 can be conditionally overexpressed under the control of the endogenous Rosa26 promoter in a hematopoietic cell-specific fashion (Tie2-Cre;R26Stop(FL)Bmi1). Although overexpression of Bmi1 did not significantly affect steady state hematopoiesis, it promoted expansion of functional HSCs during ex vivo culture and efficiently protected HSCs against loss of self-renewal capacity during serial transplantation. Overexpression of Bmi1 had no effect on DNA damage response triggered by ionizing radiation. In contrast, Tie2-Cre;R26Stop(FL)Bmi1 HSCs under oxidative stress maintained a multipotent state and generally tolerated oxidative stress better than the control. Unexpectedly, overexpression of Bmi1 had no impact on the level of intracellular reactive oxygen species (ROS).
Conclusions/significance: Our findings demonstrate that overexpression of Bmi1 confers resistance to stresses, particularly oxidative stress, onto HSCs. This thereby enhances their regenerative capacity and suggests that Bmi1 is located downstream of ROS signaling and negatively regulated by it.
Conflict of interest statement
Figures
Similar articles
-
Reciprocal expression of Bmi1 and Mel-18 is associated with functioning of primitive hematopoietic cells.Exp Hematol. 2009 Jul;37(7):857-866.e2. doi: 10.1016/j.exphem.2009.04.011. Epub 2009 May 3. Exp Hematol. 2009. PMID: 19409954
-
Poised lineage specification in multipotential hematopoietic stem and progenitor cells by the polycomb protein Bmi1.Cell Stem Cell. 2010 Mar 5;6(3):279-86. doi: 10.1016/j.stem.2010.01.005. Cell Stem Cell. 2010. PMID: 20207230
-
Long-term maintenance of human hematopoietic stem/progenitor cells by expression of BMI1.Blood. 2008 Mar 1;111(5):2621-30. doi: 10.1182/blood-2007-08-106666. Epub 2007 Dec 21. Blood. 2008. PMID: 18156489
-
Role of the polycomb group gene BMI1 in normal and leukemic hematopoietic stem and progenitor cells.Curr Opin Hematol. 2010 Jul;17(4):294-9. doi: 10.1097/MOH.0b013e328338c439. Curr Opin Hematol. 2010. PMID: 20308890 Review.
-
A protein synthesis brake for hematopoietic stem cell maintenance.Genes Dev. 2022 Aug 1;36(15-16):871-873. doi: 10.1101/gad.350107.122. Genes Dev. 2022. PMID: 36207141 Free PMC article. Review.
Cited by
-
Maternal high-fat diet and obesity compromise fetal hematopoiesis.Mol Metab. 2014 Nov 18;4(1):25-38. doi: 10.1016/j.molmet.2014.11.001. eCollection 2015 Jan. Mol Metab. 2014. PMID: 25685687 Free PMC article.
-
Role of Bmi-1 in regulation of ionizing irradiation-induced epithelial-mesenchymal transition and migration of breast cancer cells.PLoS One. 2015 Mar 3;10(3):e0118799. doi: 10.1371/journal.pone.0118799. eCollection 2015. PLoS One. 2015. PMID: 25734775 Free PMC article.
-
Role of epigenetic in cancer biology, in hematologic malignancies and in anticancer therapy.Front Mol Med. 2024 Sep 6;4:1426454. doi: 10.3389/fmmed.2024.1426454. eCollection 2024. Front Mol Med. 2024. PMID: 39308891 Free PMC article. Review.
-
Heterozygous knockout of the Bmi-1 gene causes an early onset of phenotypes associated with brain aging.Age (Dordr). 2014 Feb;36(1):129-39. doi: 10.1007/s11357-013-9552-9. Epub 2013 Jun 15. Age (Dordr). 2014. PMID: 23771506 Free PMC article.
-
Doxycycline-induced exogenous Bmi-1 expression enhances tumor formation in a murine model of oral squamous cell carcinoma.Cancer Biol Ther. 2020 May 3;21(5):400-411. doi: 10.1080/15384047.2020.1720485. Epub 2020 Feb 9. Cancer Biol Ther. 2020. PMID: 32037955 Free PMC article.
References
-
- Simon JA, Kingston RE. Mechanisms of polycomb gene silencing: knowns and unknowns. Nat Rev Mol Cell Biol. 2009;10:697–708. - PubMed
-
- Iwama A, Oguro H, Negishi M, Kato Y, Nakauchi H. Epigenetic regulation of hematopoietic stem cell self-renewal by polycomb group genes. Int J Hematol. 2005;81:294–300. - PubMed
-
- Konuma T, Oguro H, Iwama A. Role of the polycomb group proteins in hematopoietic stem cells. Dev Growth Differ. 2010;52:505–516. - PubMed
-
- Lessard J, Sauvageau G. Bmi-1 determines the proliferative capacity of normal and leukaemic stem cells. Nature. 2003;423:255–260. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
