Demasculinization of the Anopheles gambiae X chromosome
- PMID: 22607633
- PMCID: PMC3428665
- DOI: 10.1186/1471-2148-12-69
Demasculinization of the Anopheles gambiae X chromosome
Abstract
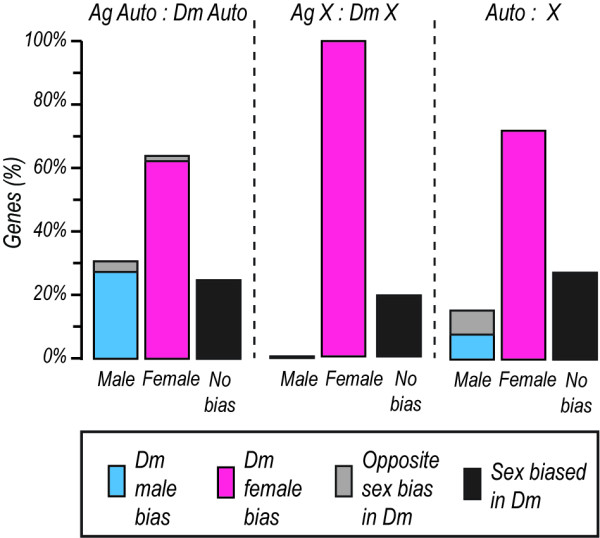
Background: In a number of organisms sex-biased genes are non-randomly distributed between autosomes and the shared sex chromosome X (or Z). Studies on Anopheles gambiae have produced conflicting results regarding the underrepresentation of male-biased genes on the X chromosome and it is unclear to what extent sexual antagonism, dosage compensation or X-inactivation in the male germline, the evolutionary forces that have been suggested to affect the chromosomal distribution of sex-biased genes, are operational in Anopheles.
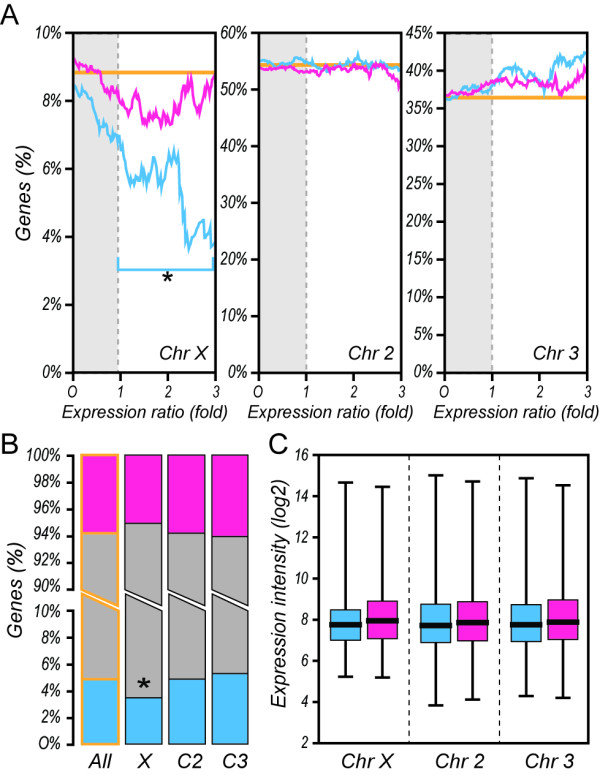
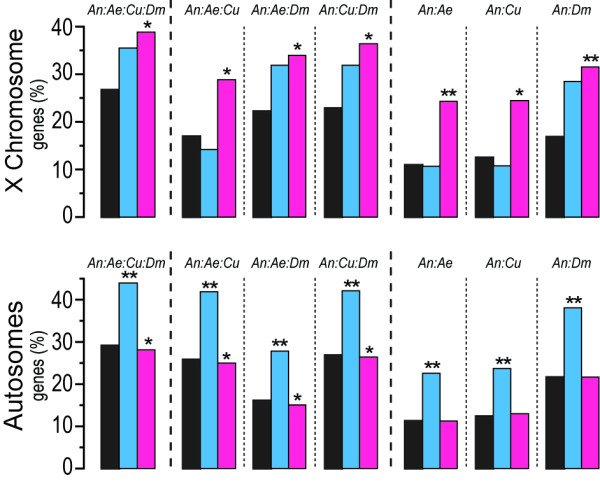
Results: We performed a meta-analysis of sex-biased gene expression in Anopheles gambiae which provides evidence for a general underrepresentation of male-biased genes on the X-chromosome that increased in significance with the observed degree of sex-bias. A phylogenomic comparison between Drosophila melanogaster, Aedes aegypti and Culex quinquefasciatus also indicates that the Anopheles X chromosome strongly disfavours the evolutionary conservation of male-biased expression and that novel male-biased genes are more likely to arise on autosomes. Finally, we demonstrate experimentally that transgenes situated on the Anopheles gambiae X chromosome are transcriptionally silenced in the male germline.
Conclusion: The data presented here support the hypothesis that the observed demasculinization of the Anopheles X chromosome is driven by X-chromosome inactivation in the male germline and by sexual antagonism. The demasculinization appears to be the consequence of a loss of male-biased expression, rather than a failure in the establishment or the extinction of male-biased genes.
Figures