

Vigilin interacts with signal peptide peptidase
- PMID: 22607704
- PMCID: PMC3490818
- DOI: 10.1186/1477-5956-10-33
Vigilin interacts with signal peptide peptidase
Abstract
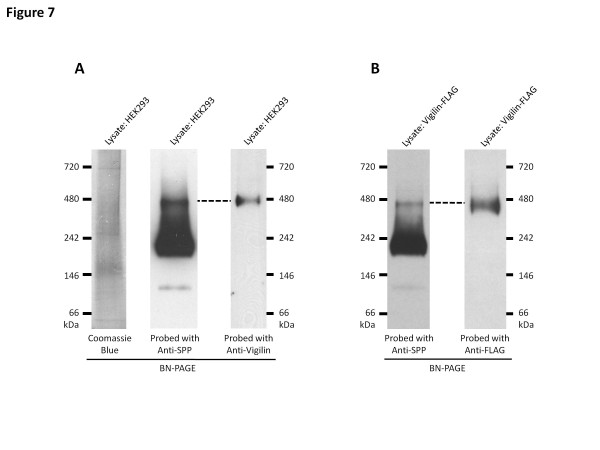
Background: Signal peptide peptidase (SPP), a member of the presenilin-like intra-membrane cleaving aspartyl protease family, migrates on Blue Native (BN) gels as 100 kDa, 200 kDa and 450 kDa species. SPP has recently been implicated in other non-proteolytic functions such as retro-translocation of MHC Class I molecules and binding of misfolded proteins in the endoplasmic reticulum (ER). These high molecular weight SPP complexes might contain additional proteins that regulate the proteolytic activity of SPP or support its non-catalytic functions.
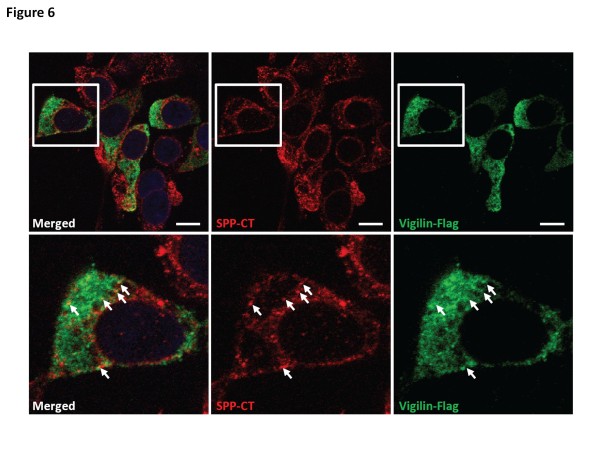
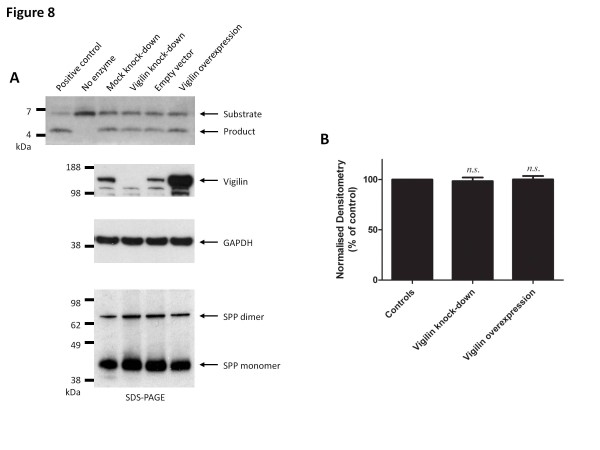
Results: In this study, an unbiased iTRAQ-labeling mass spectrometry approach was used to identify SPP-interacting proteins. We found that vigilin, a ubiquitous multi-KH domain containing cytoplasmic protein involved in RNA binding and protein translation control, selectively enriched with SPP. Vigilin interacted with SPP and both proteins co-localized in restricted intracellular domains near the ER, biochemically co-fractionated and were part of the same 450 kDa complex on BN gels. However, vigilin does not alter the protease activity of SPP, suggesting that the SPP-vigilin interaction might be involved in the non-proteolytic functions of SPP.
Conclusions: We have identified and validated vigilin as a novel interacting partner of SPP that could play an important role in the non-proteolytic functions of SPP. This data adds further weight to the idea that intramembrane-cleaving aspartyl proteases, such as presenilin and SPPs, could have other functions besides the proteolysis of short membrane stubs.
Figures

Similar articles
-
Signal peptide peptidase (SPP) assembles with substrates and misfolded membrane proteins into distinct oligomeric complexes.Biochem J. 2010 Apr 14;427(3):523-34. doi: 10.1042/BJ20091005. Biochem J. 2010. PMID: 20196774 Free PMC article.
-
Signaling Functions of Intramembrane Aspartyl-Proteases.Front Cardiovasc Med. 2020 Dec 14;7:591787. doi: 10.3389/fcvm.2020.591787. eCollection 2020. Front Cardiovasc Med. 2020. PMID: 33381526 Free PMC article. Review.
-
Differential localization and identification of a critical aspartate suggest non-redundant proteolytic functions of the presenilin homologues SPPL2b and SPPL3.J Biol Chem. 2005 Nov 25;280(47):39515-23. doi: 10.1074/jbc.M501645200. Epub 2005 Jul 5. J Biol Chem. 2005. PMID: 15998642
-
Mechanism, specificity, and physiology of signal peptide peptidase (SPP) and SPP-like proteases.Biochim Biophys Acta. 2013 Dec;1828(12):2828-39. doi: 10.1016/j.bbamem.2013.03.033. Biochim Biophys Acta. 2013. PMID: 24099004 Review.
-
Toward the structure of presenilin/γ-secretase and presenilin homologs.Biochim Biophys Acta. 2013 Dec;1828(12):2886-97. doi: 10.1016/j.bbamem.2013.04.015. Biochim Biophys Acta. 2013. PMID: 24099007 Free PMC article. Review.
Cited by
-
Structural biology of presenilin 1 complexes.Mol Neurodegener. 2014 Dec 18;9:59. doi: 10.1186/1750-1326-9-59. Mol Neurodegener. 2014. PMID: 25523933 Free PMC article. Review.
-
Structural and Chemical Biology of Presenilin Complexes.Cold Spring Harb Perspect Med. 2017 Dec 1;7(12):a024067. doi: 10.1101/cshperspect.a024067. Cold Spring Harb Perspect Med. 2017. PMID: 28320827 Free PMC article. Review.
References
-
- Weihofen A, Lemberg MK, Ploegh HL, Bogyo M, Martoglio B. Release of signal peptide fragments into the cytosol requires cleavage in the transmembrane region by a protease activity that is specifically blocked by a novel cysteine protease inhibitor. J Biol Chem. 2000;275:30951–30956. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
