

Dendritic cells in antifungal immunity and vaccine design
- PMID: 22607797
- PMCID: PMC3401965
- DOI: 10.1016/j.chom.2012.04.005
Dendritic cells in antifungal immunity and vaccine design
Abstract
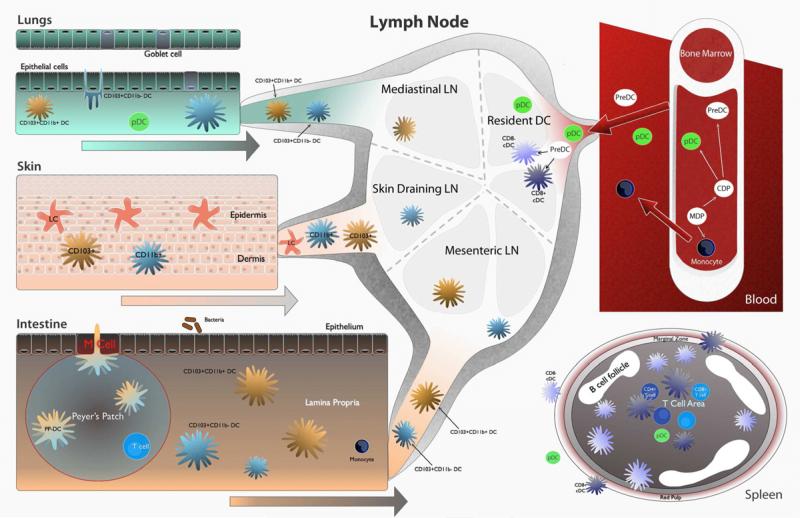
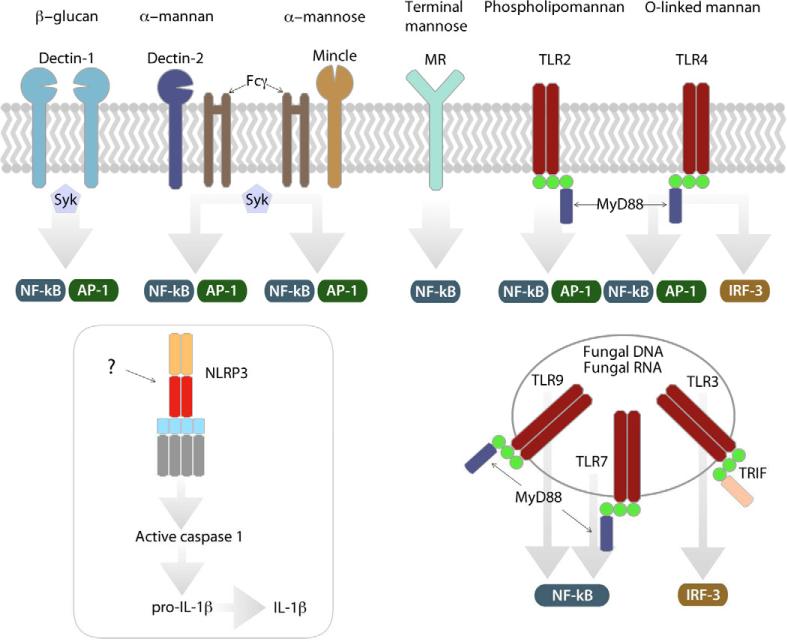
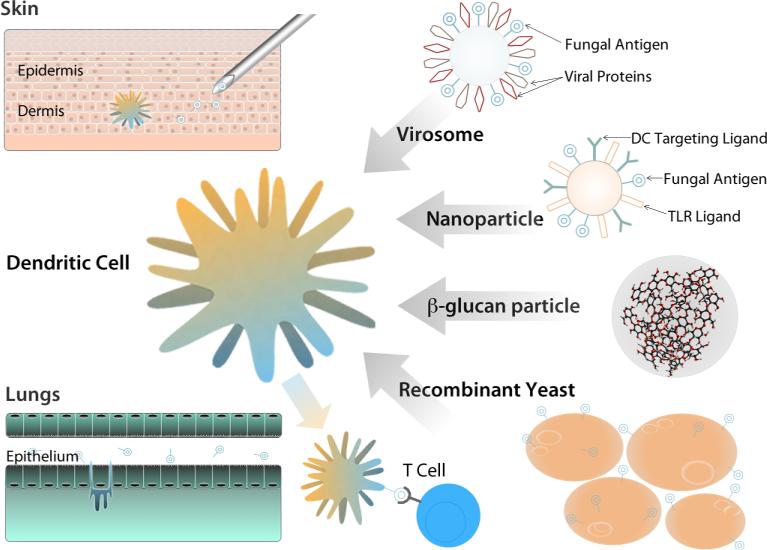
Life-threatening fungal infections have increased in recent years while treatment options remain limited. The development of vaccines against fungal pathogens represents a key advance sorely needed to combat the increasing fungal disease threat. Dendritic cells (DC) are uniquely able to shape antifungal immunity by initiating and modulating naive T cell responses. Targeting DC may allow for the generation of potent vaccines against fungal pathogens. In the context of antifungal vaccine design, we describe the characteristics of the varied DC subsets, how DC recognize fungi, their function in immunity against fungal pathogens, and how DC can be targeted in order to create new antifungal vaccines. Ongoing studies continue to highlight the critical role of DC in antifungal immunity and will help guide DC-based vaccine strategies.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Exciting developments in the immunology of fungal infections.Cell Host Microbe. 2012 May 17;11(5):422-4. doi: 10.1016/j.chom.2012.04.010. Cell Host Microbe. 2012. PMID: 22607795
References
-
- Abdel-Motal UM, Berg L, Rosen A, Bengtsson M, Thorpe CJ, Kihlberg J, Dahmen J, Magnusson G, Karlsson KA, Jondal M. Immunization with glycosylated Kb-binding peptides generates carbohydrate-specific, unrestricted cytotoxic T cells. Eur J Immunol. 1996;26:544–551. - PubMed
-
- Allan RS, Waithman J, Bedoui S, Jones CM, Villadangos JA, Zhan Y, Lew AM, Shortman K, Heath WR, Carbone FR. Migratory dendritic cells transfer antigen to a lymph node-resident dendritic cell population for efficient CTL priming. Immunity. 2006;25:153–162. - PubMed
-
- Backer R, van Leeuwen F, Kraal G, den Haan JM. CD8- dendritic cells preferentially cross-present Saccharomyces cerevisiae antigens. Eur J Immunol. 2008;38:370–380. - PubMed
-
- Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
