

Disruption of Fnip1 reveals a metabolic checkpoint controlling B lymphocyte development
- PMID: 22608497
- PMCID: PMC3361584
- DOI: 10.1016/j.immuni.2012.02.019
Disruption of Fnip1 reveals a metabolic checkpoint controlling B lymphocyte development
Abstract
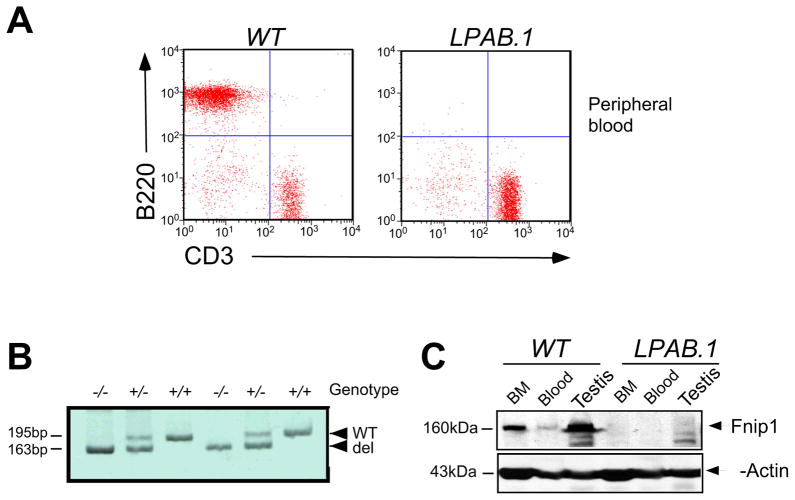
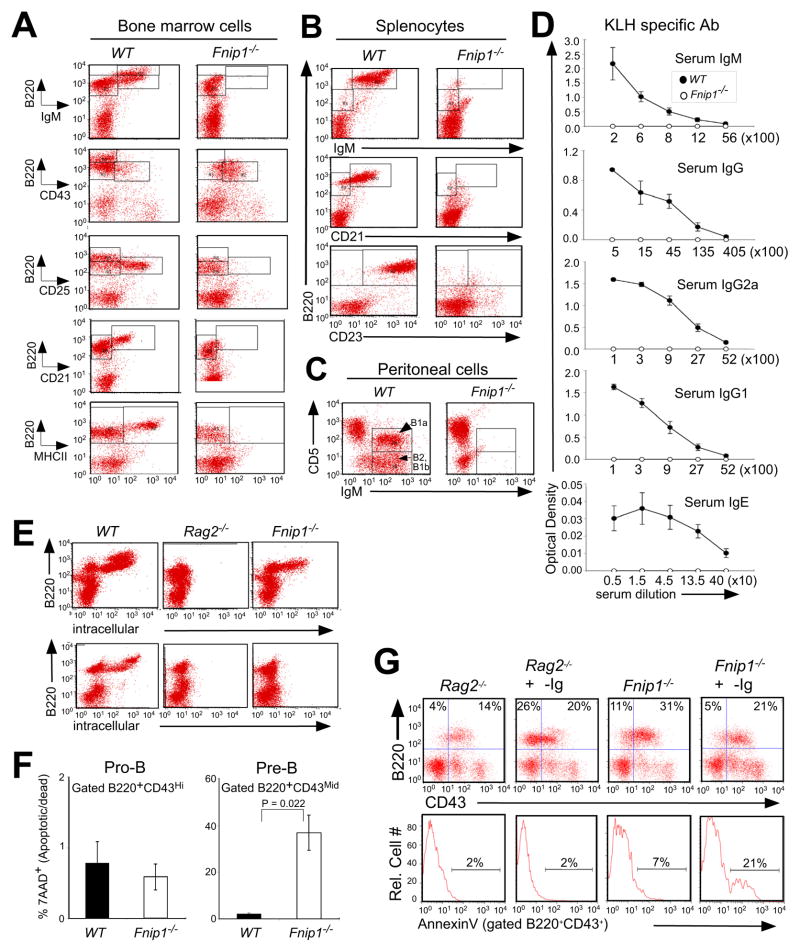
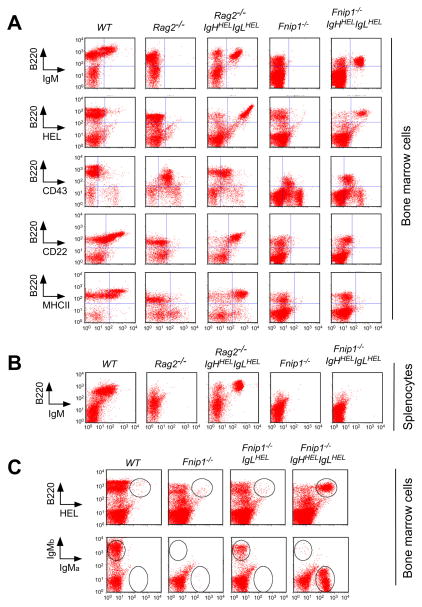
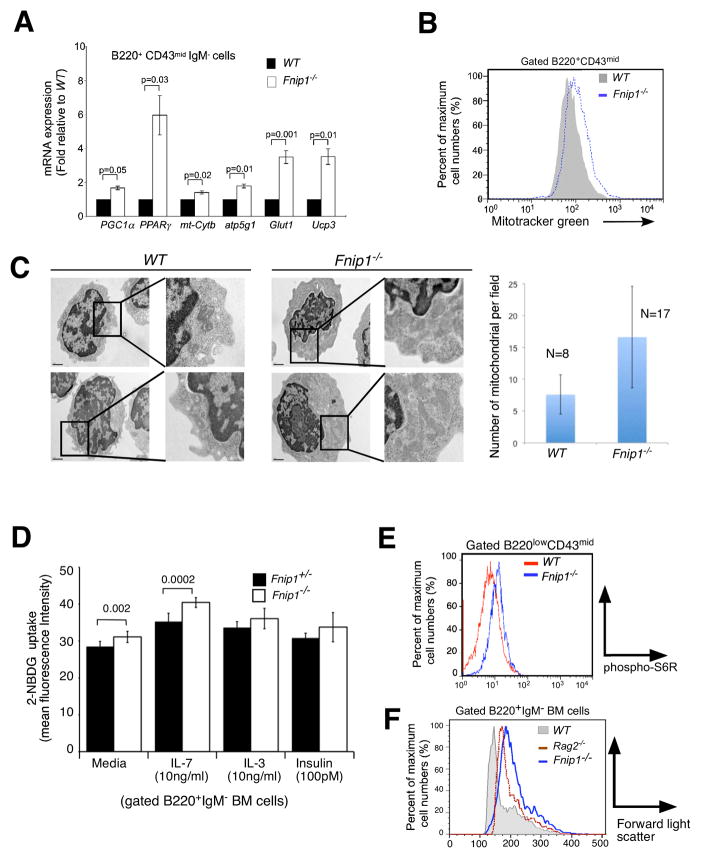
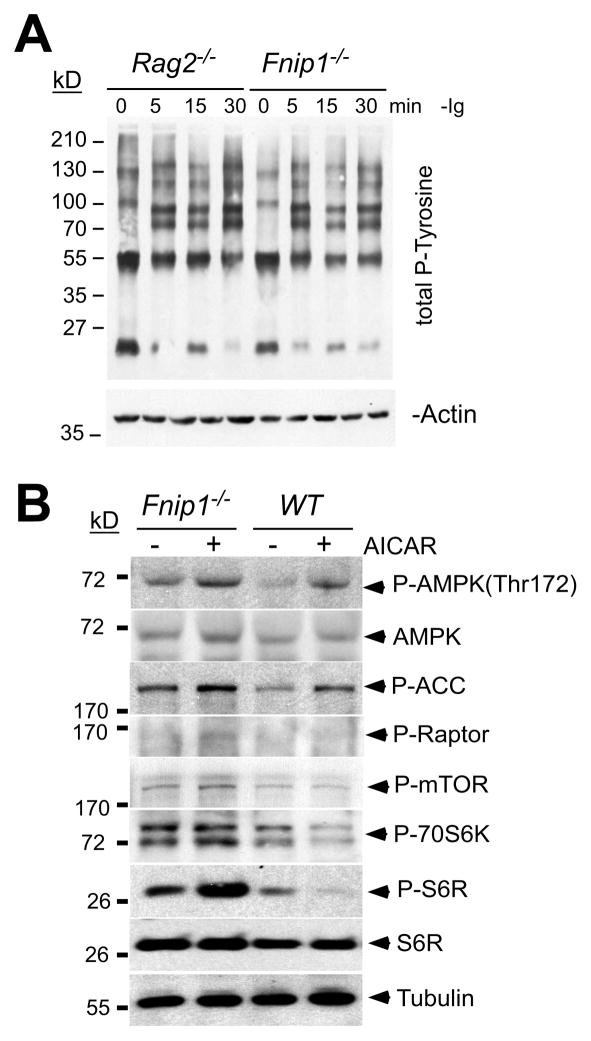
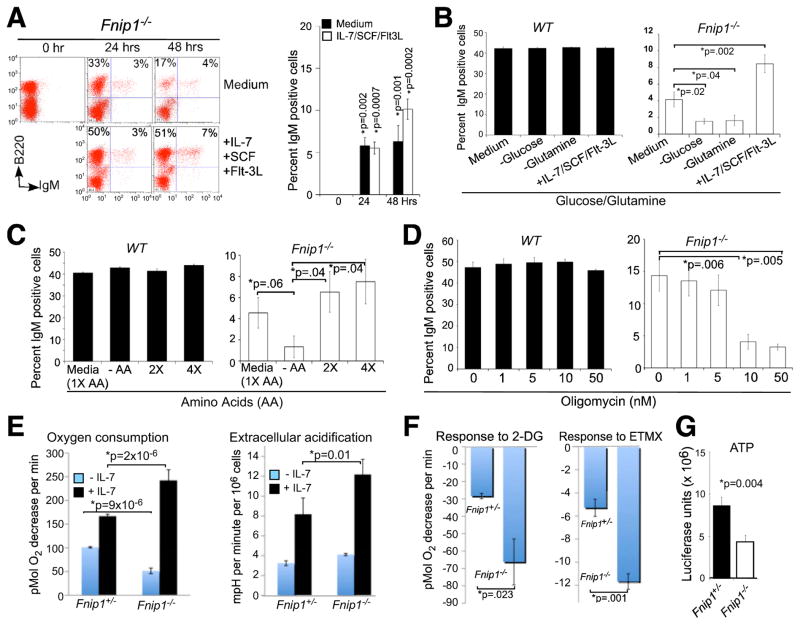
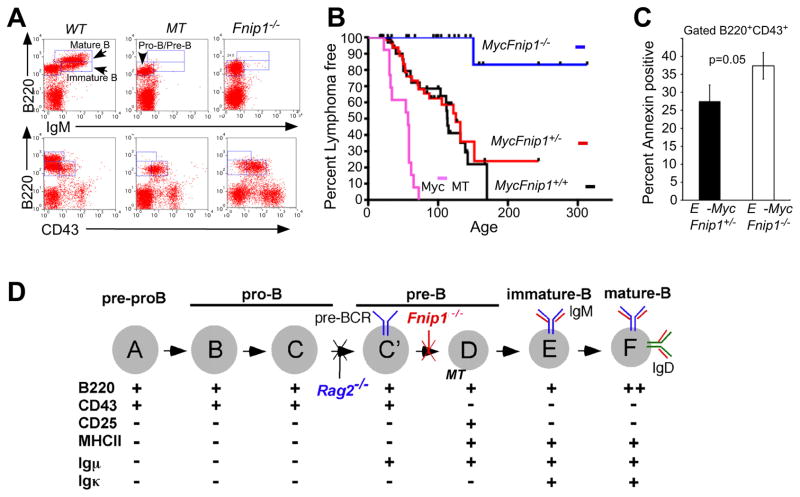
The coordination of nutrient and energy availability with cell growth and division is essential for proper immune cell development and function. By using a chemical mutagenesis strategy in mice, we identified a pedigree that has a complete block in B cell development at the pre-B cell stage resulting from a deletion in the Fnip1 gene. Enforced expression of an immunoglobulin transgene failed to rescue B cell development. Whereas essential pre-B cell signaling molecules were activated normally in Fnip1-null pre-B cells, the metabolic regulators AMPK and mTOR were dysregulated, resulting in excessive cell growth and enhanced sensitivity to apoptosis in response to metabolic stress (pre-B cell receptor crosslinking, oncogene activation). These results indicate that Folliculin-interacting protein 1 (Fnip1) is vital for B cell development and metabolic homeostasis and reveal a metabolic checkpoint that may ensure that pre-B cells have sufficient metabolic capacity to support division, while limiting lymphomagenesis caused by deregulated growth.
Copyright © 2012 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors have no competing financial interests.
Figures
Similar articles
-
Metabolic regulator Fnip1 is crucial for iNKT lymphocyte development.Proc Natl Acad Sci U S A. 2014 May 13;111(19):7066-71. doi: 10.1073/pnas.1406473111. Epub 2014 Apr 30. Proc Natl Acad Sci U S A. 2014. PMID: 24785297 Free PMC article.
-
Folliculin Interacting Protein 1 Maintains Metabolic Homeostasis during B Cell Development by Modulating AMPK, mTORC1, and TFE3.J Immunol. 2019 Dec 1;203(11):2899-2908. doi: 10.4049/jimmunol.1900395. Epub 2019 Nov 1. J Immunol. 2019. PMID: 31676673 Free PMC article.
-
Control of B lymphocyte development and functions by the mTOR signaling pathways.Cytokine Growth Factor Rev. 2017 Jun;35:47-62. doi: 10.1016/j.cytogfr.2017.04.005. Epub 2017 May 22. Cytokine Growth Factor Rev. 2017. PMID: 28583723 Free PMC article. Review.
-
Folliculin encoded by the BHD gene interacts with a binding protein, FNIP1, and AMPK, and is involved in AMPK and mTOR signaling.Proc Natl Acad Sci U S A. 2006 Oct 17;103(42):15552-7. doi: 10.1073/pnas.0603781103. Epub 2006 Oct 6. Proc Natl Acad Sci U S A. 2006. PMID: 17028174 Free PMC article.
-
The PI3K pathway in B cell metabolism.Crit Rev Biochem Mol Biol. 2016 Sep;51(5):359-378. doi: 10.1080/10409238.2016.1215288. Epub 2016 Aug 5. Crit Rev Biochem Mol Biol. 2016. PMID: 27494162 Free PMC article. Review.
Cited by
-
Familial multiple discoid fibromas is linked to a locus on chromosome 5 including the FNIP1 gene.J Hum Genet. 2023 Apr;68(4):273-279. doi: 10.1038/s10038-022-01113-1. Epub 2023 Jan 4. J Hum Genet. 2023. PMID: 36599954
-
Loss of Fnip1 alters kidney developmental transcriptional program and synergizes with TSC1 loss to promote mTORC1 activation and renal cyst formation.PLoS One. 2018 Jun 13;13(6):e0197973. doi: 10.1371/journal.pone.0197973. eCollection 2018. PLoS One. 2018. PMID: 29897930 Free PMC article.
-
Metabolic regulator Fnip1 is crucial for iNKT lymphocyte development.Proc Natl Acad Sci U S A. 2014 May 13;111(19):7066-71. doi: 10.1073/pnas.1406473111. Epub 2014 Apr 30. Proc Natl Acad Sci U S A. 2014. PMID: 24785297 Free PMC article.
-
A defined metabolic state in pre B cells governs B-cell development and is counterbalanced by Swiprosin-2/EFhd1.Cell Death Differ. 2017 Jul;24(7):1239-1252. doi: 10.1038/cdd.2017.52. Epub 2017 May 19. Cell Death Differ. 2017. PMID: 28524857 Free PMC article.
-
Absent B cells, agammaglobulinemia, and hypertrophic cardiomyopathy in folliculin-interacting protein 1 deficiency.Blood. 2021 Jan 28;137(4):493-499. doi: 10.1182/blood.2020006441. Blood. 2021. PMID: 32905580 Free PMC article.
References
-
- Adams JM, Harris AW, Pinkert CA, Corcoran LM, Alexander WS, Cory S, Palmiter RD, Brinster RL. The c-myc oncogene driven by immunoglobulin enhancers induces lymphoid malignancy in transgenic mice. Nature. 1985;318:533–538. - PubMed
-
- Allen CD, Okada T, Tang HL, Cyster JG. Imaging of germinal center selection events during affinity maturation. Science. 2007;315:528–531. - PubMed
-
- Appleby MW, Ramsdell F. A forward-genetic approach for analysis of the immune system. Nat Rev Immunol. 2003;3:463–471. - PubMed
-
- Arad M, Seidman CE, Seidman JG. AMP-activated protein kinase in the heart: role during health and disease. Circ Res. 2007;100:474–488. - PubMed
-
- Baba M, Hong SB, Sharma N, Warren MB, Nickerson ML, Iwamatsu A, Esposito D, Gillette WK, Hopkins RF, 3rd, Hartley JL, et al. Folliculin encoded by the BHD gene interacts with a binding protein, FNIP1, and AMPK, and is involved in AMPK and mTOR signaling. Proc Natl Acad Sci U S A. 2006;103:15552–15557. - PMC - PubMed
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
