

The organochlorine o,p'-DDT plays a role in coactivator-mediated MAPK crosstalk in MCF-7 breast cancer cells
- PMID: 22609851
- PMCID: PMC3440107
- DOI: 10.1289/ehp.1104296
The organochlorine o,p'-DDT plays a role in coactivator-mediated MAPK crosstalk in MCF-7 breast cancer cells
Abstract
Background: The organochlorine dichlorodiphenyltrichloroethane (DDT), a known estrogen mimic and endocrine disruptor, has been linked to animal and human disorders. However, the detailed mechanism(s) by which DDT affects cellular physiology remains incompletely defined.
Objectives: We and others have shown that DDT activates cell-signaling cascades, culminating in the activation of estrogen receptor-dependent and -independent gene expression. Here, we identify a mechanism by which DDT alters cellular signaling and gene expression, independent of the estrogen receptor.
Methods: We performed quantitative polymerase chain reaction array analysis of gene expression in MCF-7 breast cancer cells using either estradiol (E₂) or o,p´-DDT to identify distinct cellular gene expression responses. To elucidate the mechanisms by which DDT regulates cell signaling, we used molecular and pharmacological techniques.
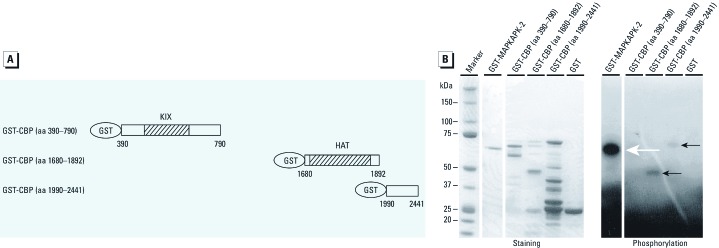
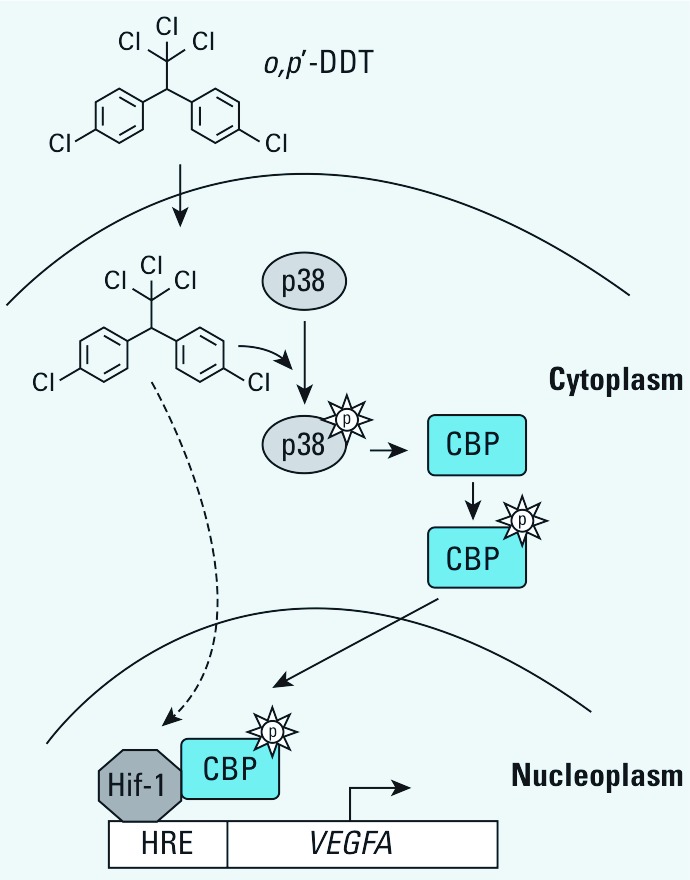
Results: E₂ and DDT treatment both altered the expression of many of the genes assayed, but up-regulation of vascular endothelial growth factor A (VEGFA) was observed only after DDT treatment, and this increase was not affected by the pure estrogen receptor α antagonist ICI 182780. Furthermore, DDT increased activation of the HIF-1 response element (HRE), a known enhancer of the VEGFA gene. This DDT-mediated increase in HRE activity was augmented by the coactivator CBP (CREB-binding protein) and was dependent on the p38 pathway.
Conclusions: DDT up-regulated the expression of several genes in MCF-7 breast cancer cells that were not altered by treatment with E₂, including VEGFA. We propose that this DDT-initiated, ER-independent stimulation of gene expression is due to DDT's ability to initiate crosstalk between MAPK (mitogen-activated protein kinase) signaling pathways and transcriptional coactivators.
Conflict of interest statement
The authors declare they have no actual or potential competing financial interests.
Figures




References
-
- Ahlborg UG, Lipworth L, Titus-Ernstoff L, Hsieh CC, Hanberg A, Baron J, et al. Organochlorine compounds in relation to breast cancer, endometrial cancer, and endometriosis: an assessment of the biological and epidemiological evidence. Crit Rev Toxicol. 1995;25(6):463–531. - PubMed
-
- Aigner EJ, Leone AD, Falconer RL. Concentrations and enantiomeric ratios of organochlorine pesticides in soils from the U.S. Corn Belt. Environ Sci Technol. 1998;32(9):1162–1168.
-
- Ait-Si-Ali S, Carlisi D, Ramirez S, Upegui-Gonzalez LC, Duquet A, Robin P, et al. Phosphorylation by p44 MAP kinase/ERK1 stimulates CBP histone acetyl transferase activity in vitro. Biochem Biophys Res Commun. 1999;262(1):157–162. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
