

Functional analyses of mycobacterial lipoprotein diacylglyceryl transferase and comparative secretome analysis of a mycobacterial lgt mutant
- PMID: 22609911
- PMCID: PMC3416517
- DOI: 10.1128/JB.00127-12
Functional analyses of mycobacterial lipoprotein diacylglyceryl transferase and comparative secretome analysis of a mycobacterial lgt mutant
Abstract
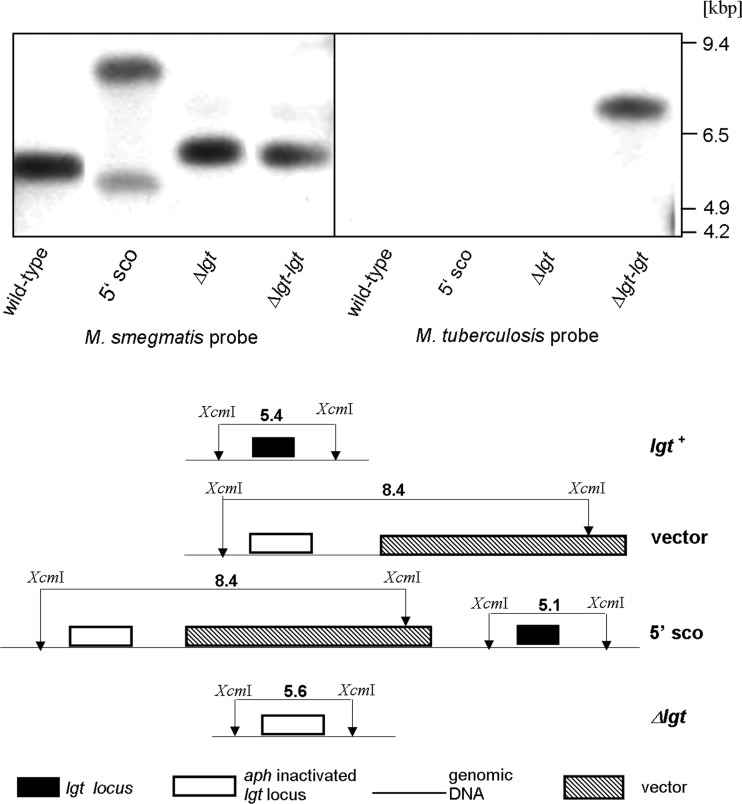
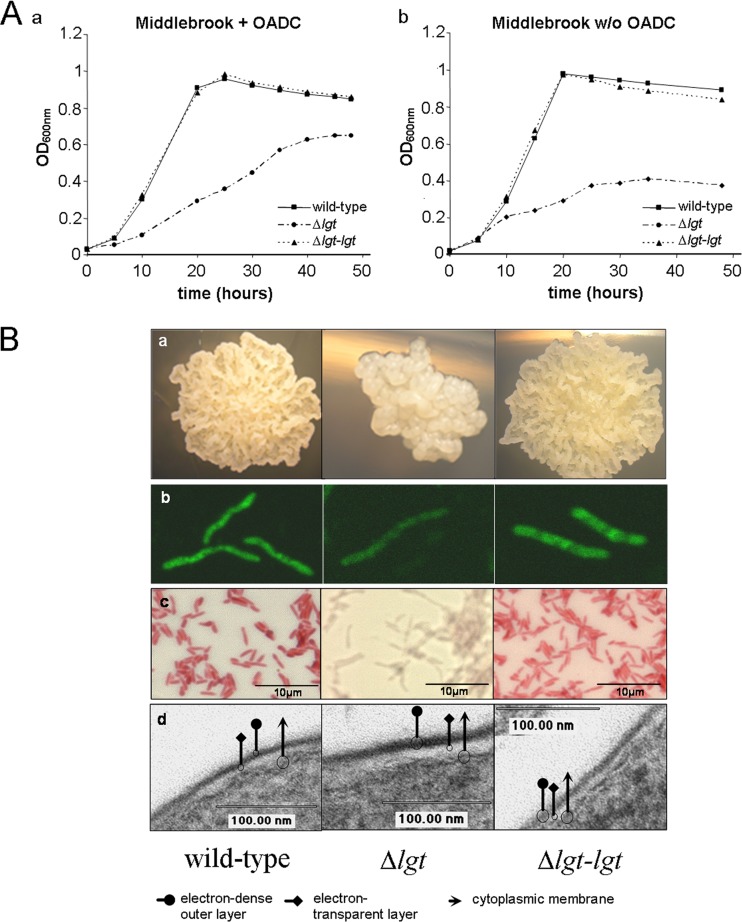
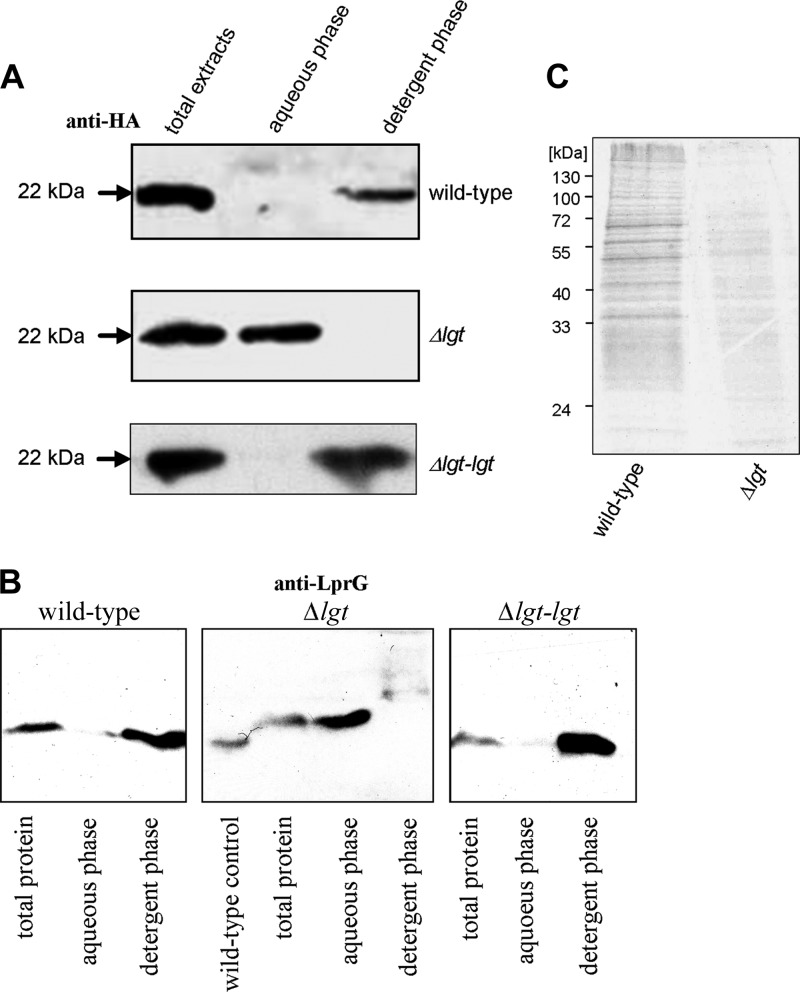
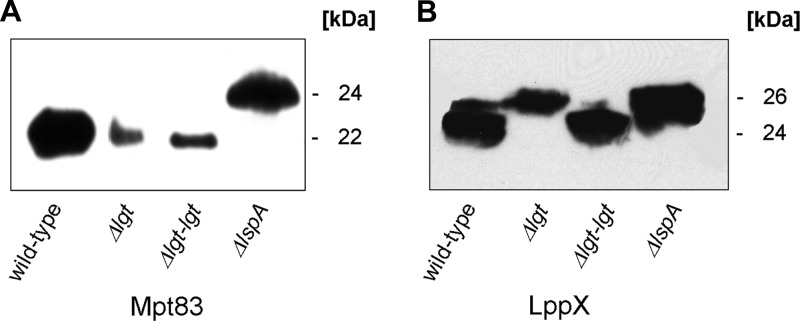
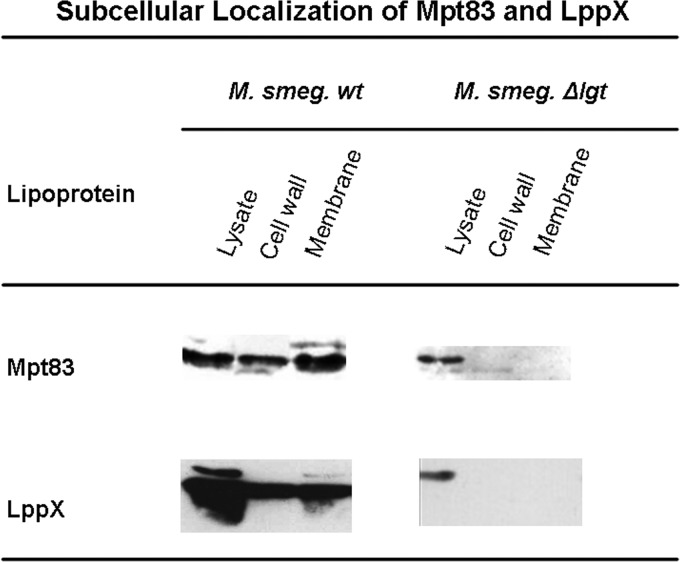
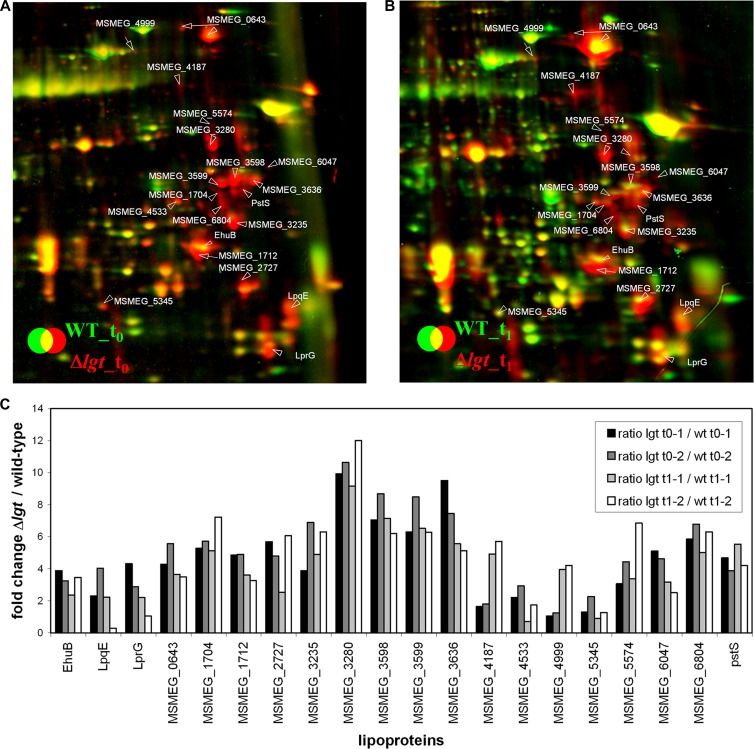
Preprolipopoprotein diacylglyceryl transferase (Lgt) is the gating enzyme of lipoprotein biosynthesis, and it attaches a lipid structure to the N-terminal part of preprolipoproteins. Using Lgt from Escherichia coli in a BLASTp search, we identified the corresponding Lgt homologue in Mycobacterium tuberculosis and two homologous (MSMEG_3222 and MSMEG_5408) Lgt in Mycobacterium smegmatis. M. tuberculosis lgt was shown to be essential, but an M. smegmatis ΔMSMEG_3222 mutant could be generated. Using Triton X-114 phase separation and [(14)C]palmitic acid incorporation, we demonstrate that MSMEG_3222 is the major Lgt in M. smegmatis. Recombinant M. tuberculosis lipoproteins Mpt83 and LppX are shown to be localized in the cell envelope of parental M. smegmatis but were absent from the cell membrane and cell wall in the M. smegmatis ΔMSMEG_3222 strain. In a proteomic study, 106 proteins were identified and quantified in the secretome of wild-type M. smegmatis, including 20 lipoproteins. All lipoproteins were secreted at higher levels in the ΔMSMEG_3222 mutant. We identify the major Lgt in M. smegmatis, show that lipoproteins lacking the lipid anchor are secreted into the culture filtrate, and demonstrate that M. tuberculosis lgt is essential and thus a validated drug target.
Figures
Similar articles
-
Mycobacterium tuberculosis Rv1302 and Mycobacterium smegmatis MSMEG_4947 have WecA function and MSMEG_4947 is required for the growth of M. smegmatis.FEMS Microbiol Lett. 2010 Sep 1;310(1):54-61. doi: 10.1111/j.1574-6968.2010.02045.x. Epub 2010 Jun 23. FEMS Microbiol Lett. 2010. PMID: 20637039
-
Correlation of over-expression of rv1900c with enhanced survival of M. smegmatis under stress conditions: Modulation of cell surface properties.Gene. 2021 Jul 30;791:145720. doi: 10.1016/j.gene.2021.145720. Epub 2021 May 18. Gene. 2021. PMID: 34019937
-
Role of the unique, non-essential phosphatidylglycerol::prolipoprotein diacylglyceryl transferase (Lgt) in Corynebacterium glutamicum.Microbiology (Reading). 2020 Aug;166(8):759-776. doi: 10.1099/mic.0.000937. Epub 2020 Jun 3. Microbiology (Reading). 2020. PMID: 32490790
-
Inhibition of Escherichia coli Lipoprotein Diacylglyceryl Transferase Is Insensitive to Resistance Caused by Deletion of Braun's Lipoprotein.J Bacteriol. 2021 Jun 8;203(13):e0014921. doi: 10.1128/JB.00149-21. Epub 2021 Jun 8. J Bacteriol. 2021. PMID: 33875545 Free PMC article.
-
Mycobacterium smegmatis: The Vanguard of Mycobacterial Research.J Bacteriol. 2023 Jan 26;205(1):e0033722. doi: 10.1128/jb.00337-22. Epub 2023 Jan 4. J Bacteriol. 2023. PMID: 36598232 Free PMC article. Review.
Cited by
-
T(H)17-Mediated Protection against Pneumococcal Carriage by a Whole-Cell Vaccine Is Dependent on Toll-Like Receptor 2 and Surface Lipoproteins.Clin Vaccine Immunol. 2015 Aug;22(8):909-16. doi: 10.1128/CVI.00118-15. Epub 2015 Jun 3. Clin Vaccine Immunol. 2015. PMID: 26041040 Free PMC article.
-
Microbial Musings - August 2020.Microbiology (Reading). 2020 Aug;166(8):680-682. doi: 10.1099/mic.0.000969. Microbiology (Reading). 2020. PMID: 32854815 Free PMC article. No abstract available.
-
PLGA particulate subunit tuberculosis vaccines promote humoral and Th17 responses but do not enhance control of Mycobacterium tuberculosis infection.PLoS One. 2018 Mar 19;13(3):e0194620. doi: 10.1371/journal.pone.0194620. eCollection 2018. PLoS One. 2018. PMID: 29554138 Free PMC article.
-
Lipoproteins of slow-growing Mycobacteria carry three fatty acids and are N-acylated by apolipoprotein N-acyltransferase BCG_2070c.BMC Microbiol. 2013 Oct 5;13:223. doi: 10.1186/1471-2180-13-223. BMC Microbiol. 2013. PMID: 24093492 Free PMC article.
-
Toward Understanding the Essence of Post-Translational Modifications for the Mycobacterium tuberculosis Immunoproteome.Front Immunol. 2014 Aug 11;5:361. doi: 10.3389/fimmu.2014.00361. eCollection 2014. Front Immunol. 2014. PMID: 25157249 Free PMC article. Review.
References
-
- Antelmann H, et al. 2001. A proteomic view on genome-based signal peptide predictions. Genome Res. 11:1484–1502 - PubMed
-
- Bradford MM. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248–254 - PubMed
-
- Brulle JK, et al. 2010. Cloning, expression and characterization of Mycobacterium tuberculosis lipoprotein LprF. Biochem. Biophys. Res. Commun. 391:679–684 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
