

Identification of a novel Wnt5a-CK1ɛ-Dvl2-Plk1-mediated primary cilia disassembly pathway
- PMID: 22609948
- PMCID: PMC3400010
- DOI: 10.1038/emboj.2012.144
Identification of a novel Wnt5a-CK1ɛ-Dvl2-Plk1-mediated primary cilia disassembly pathway
Abstract
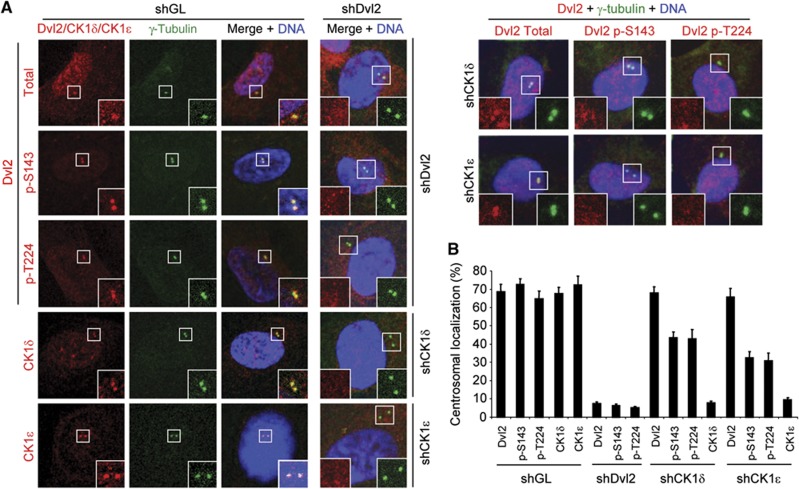
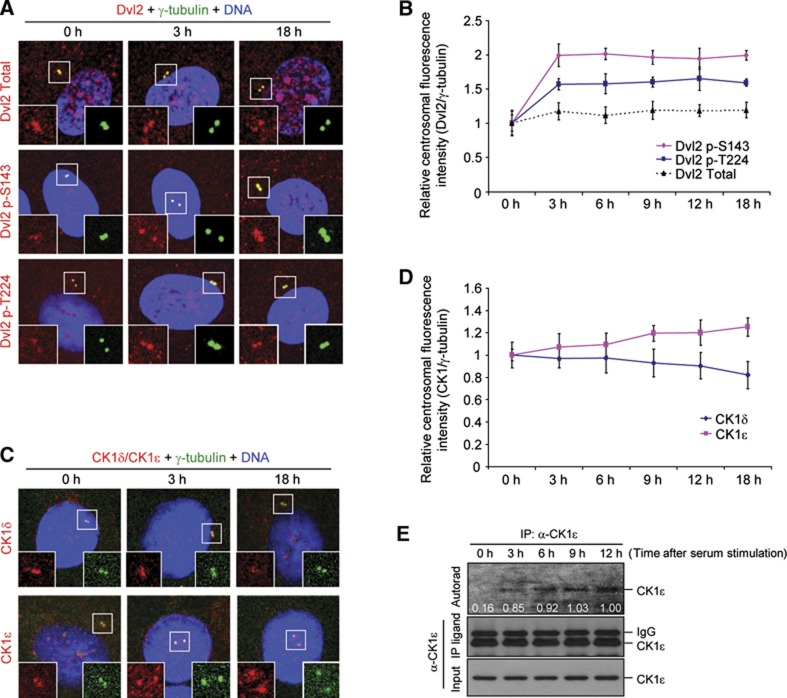
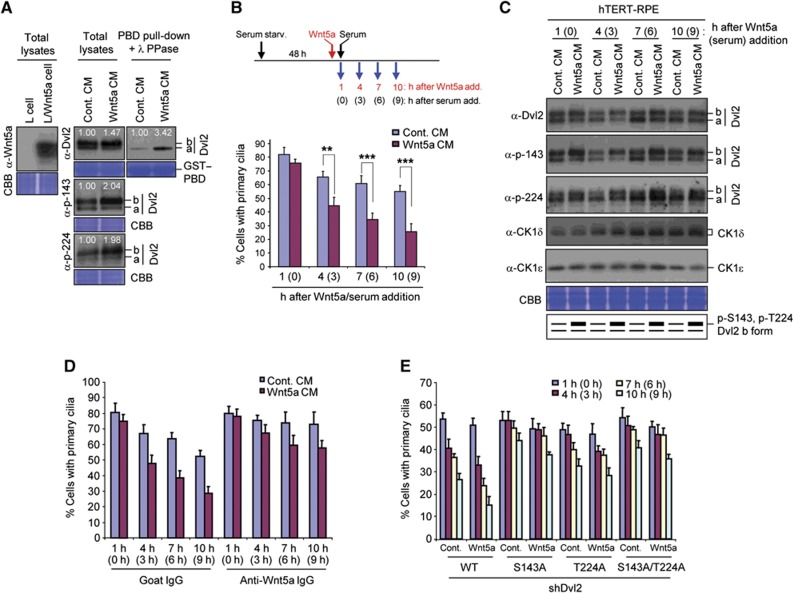
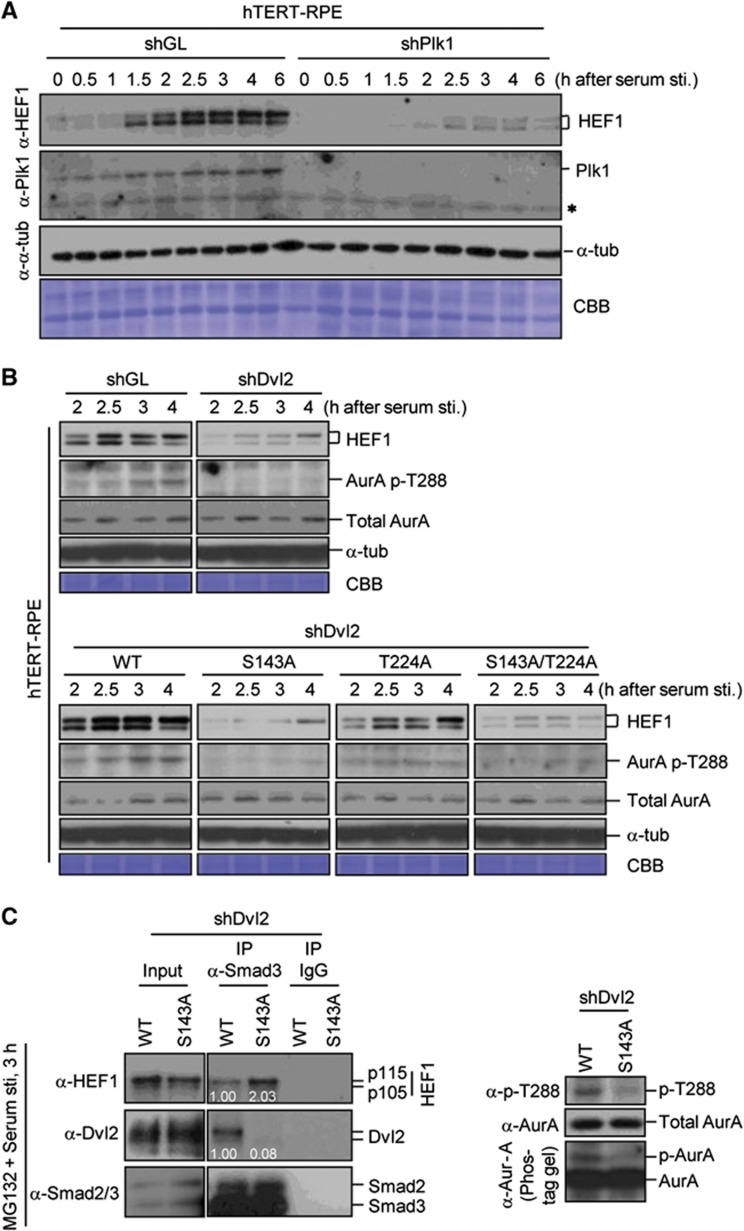
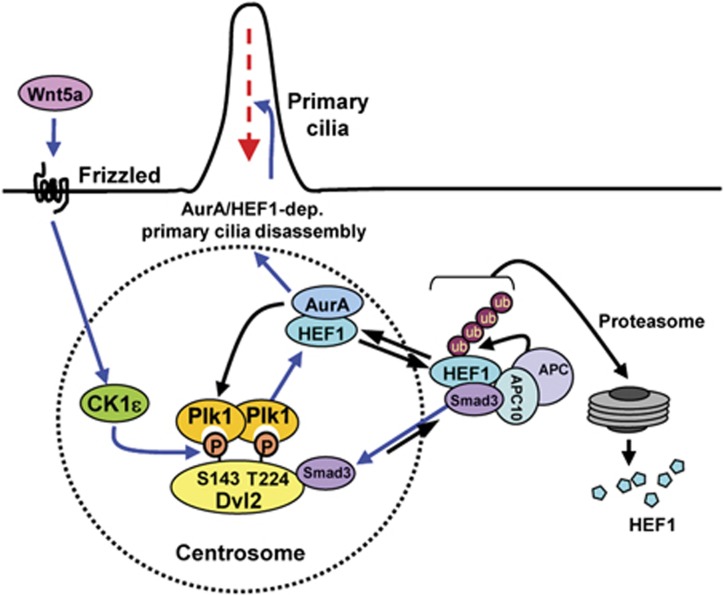
Non-motile primary cilium is an antenna-like structure whose defect is associated with a wide range of pathologies, including developmental disorders and cancer. Although mechanisms regulating cilia assembly have been extensively studied, how cilia disassembly is regulated remains poorly understood. Here, we report unexpected roles of Dishevelled 2 (Dvl2) and interphase polo-like kinase 1 (Plk1) in primary cilia disassembly. We demonstrated that Dvl2 is phosphorylated at S143 and T224 in a manner that requires both non-canonical Wnt5a ligand and casein kinase 1 epsilon (CK1ɛ), and that this event is critical to interact with Plk1 in early stages of the cell cycle. The resulting Dvl2-Plk1 complex mediated Wnt5a-CK1ɛ-Dvl2-dependent primary cilia disassembly by stabilizing the HEF1 scaffold and activating its associated Aurora-A (AurA), a kinase crucially required for primary cilia disassembly. Thus, via the formation of the Dvl2-Plk1 complex, Plk1 plays an unanticipated role in primary cilia disassembly by linking Wnt5a-induced biochemical steps to HEF1/AurA-dependent cilia disassembly. This study may provide new insights into the mechanism underlying ciliary disassembly processes and various cilia-related disorders.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
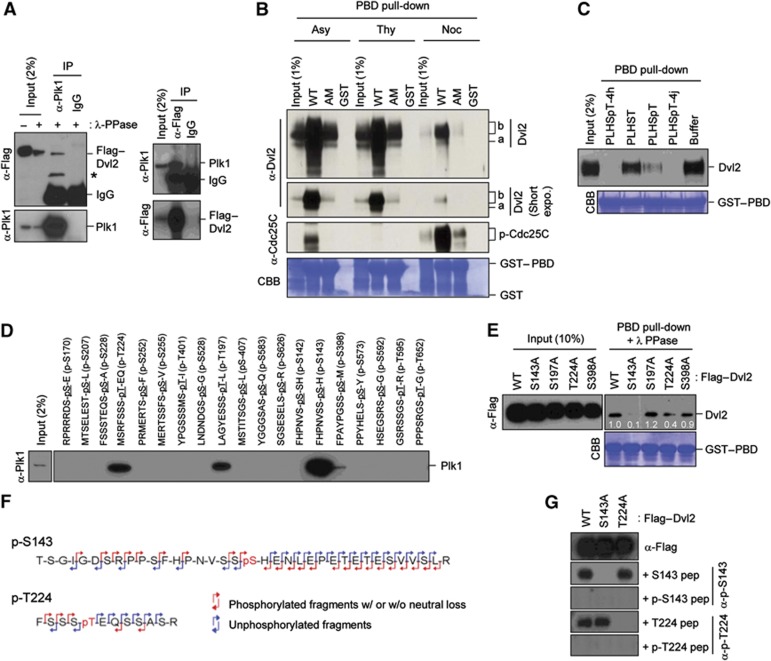
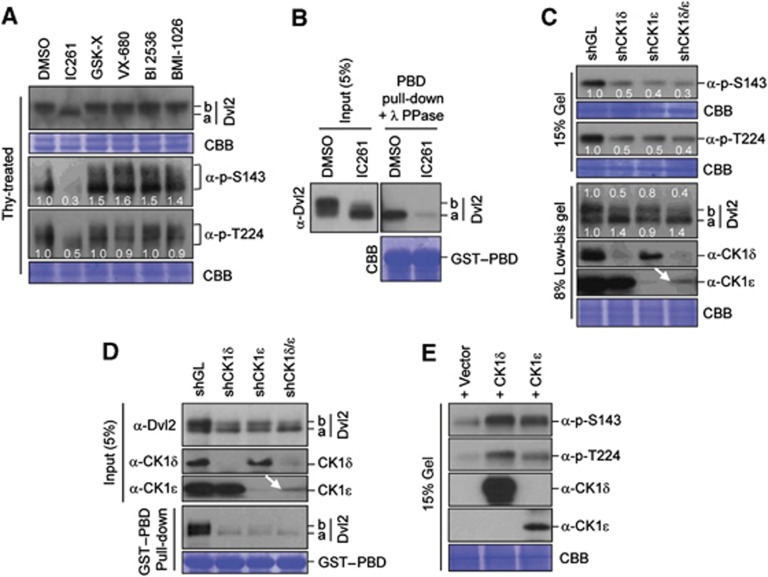
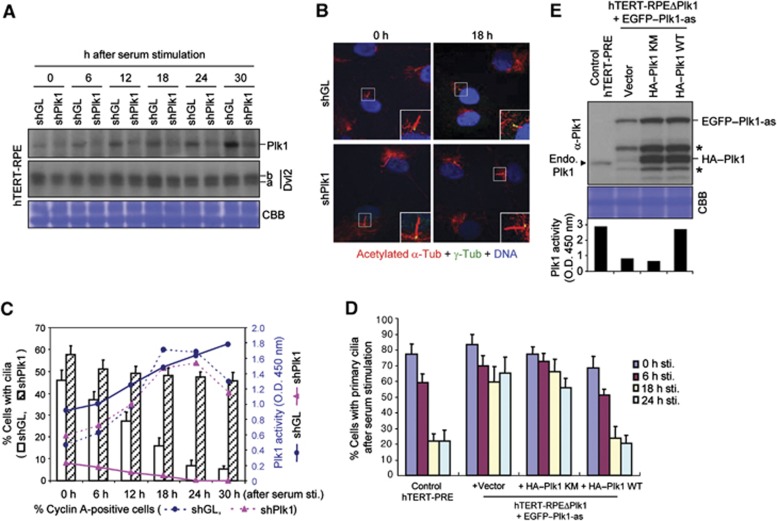
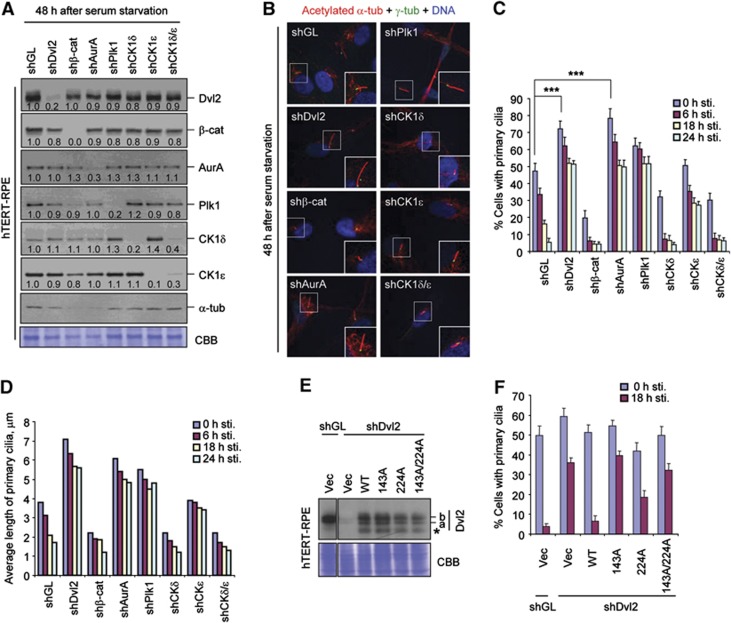
References
-
- Adams M, Smith UM, Logan CV, Johnson CA (2008) Recent advances in the molecular pathology, cell biology and genetics of ciliopathies. J Med Genet 45: 257–267 - PubMed
-
- Alieva IB, Gorgidze LA, Komarova YA, Chernobelskaya OA, Vorobjev IS (1999) Experimental model for studying the primary cilia in tissue culture cells. Membr Cell Biol 12: 895–905 - PubMed
-
- Angers S, Moon RT (2009) Proximal events in Wnt signal transduction. Nat Rev Mol Cell Biol 10: 468–477 - PubMed
-
- Archambault V, Glover DM (2009) Polo-like kinases: conservation and divergence in their functions and regulation. Nat Rev Mol Cell Biol 10: 265–275 - PubMed
-
- Barr FA, Sillje HH, Nigg EA (2004) Polo-like kinases and the orchestration of cell division. Nat Rev Mol Cell Biol 5: 429–440 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
