

Inferring gene regulatory logic from high-throughput measurements of thousands of systematically designed promoters
- PMID: 22609971
- PMCID: PMC3374032
- DOI: 10.1038/nbt.2205
Inferring gene regulatory logic from high-throughput measurements of thousands of systematically designed promoters
Abstract
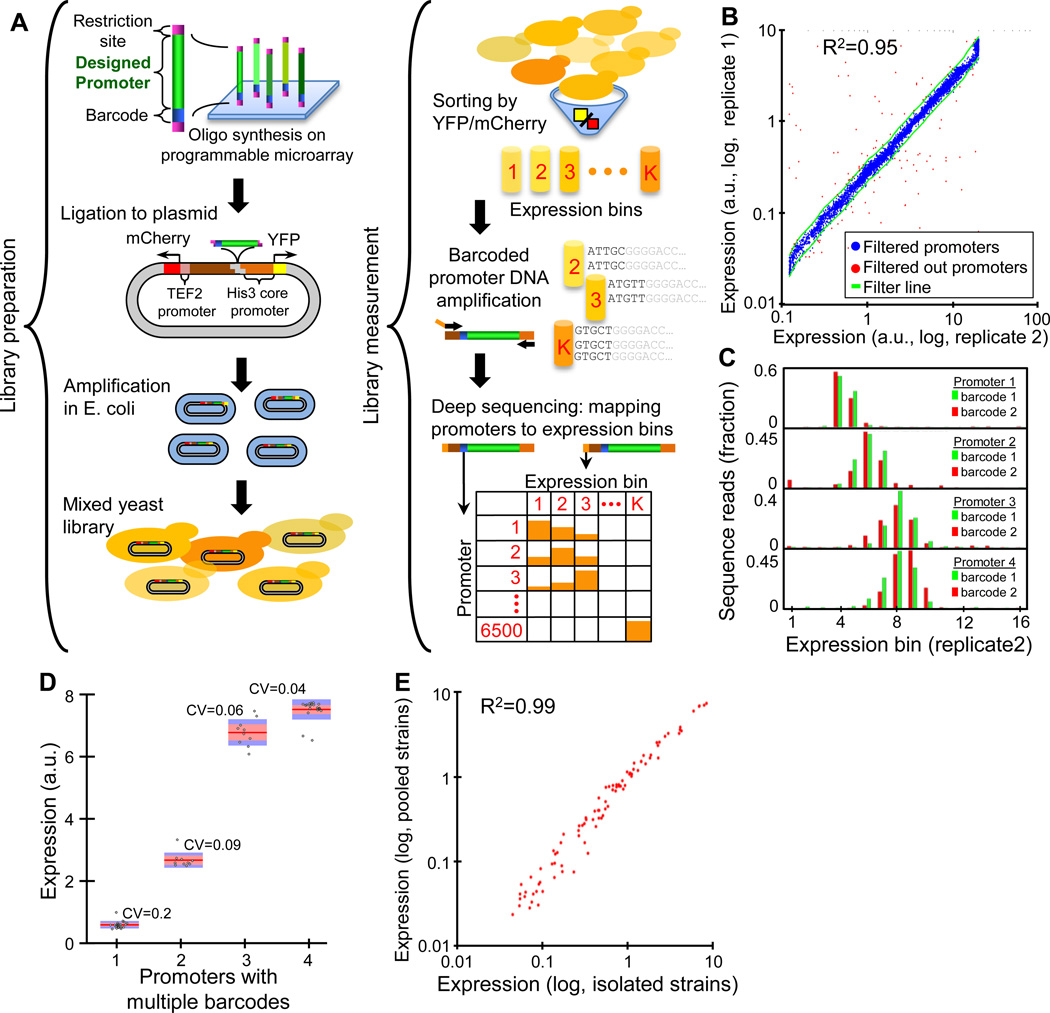
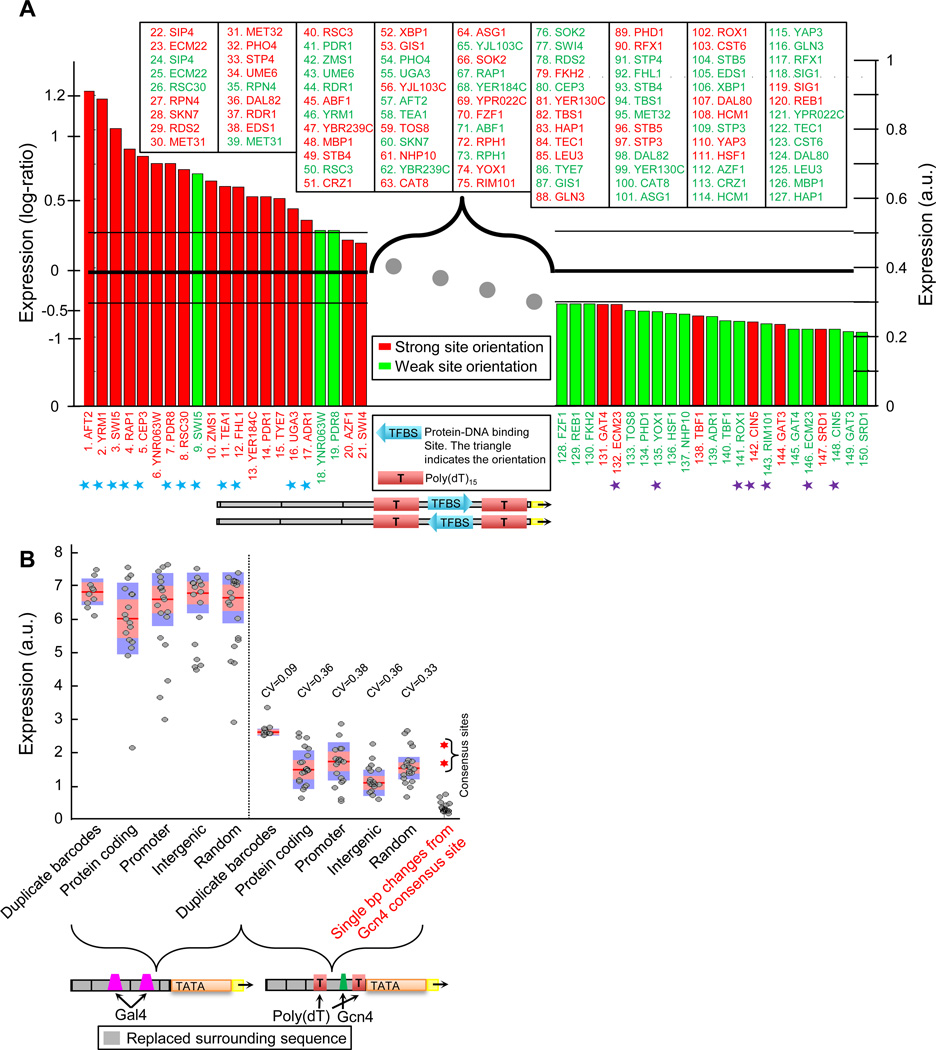
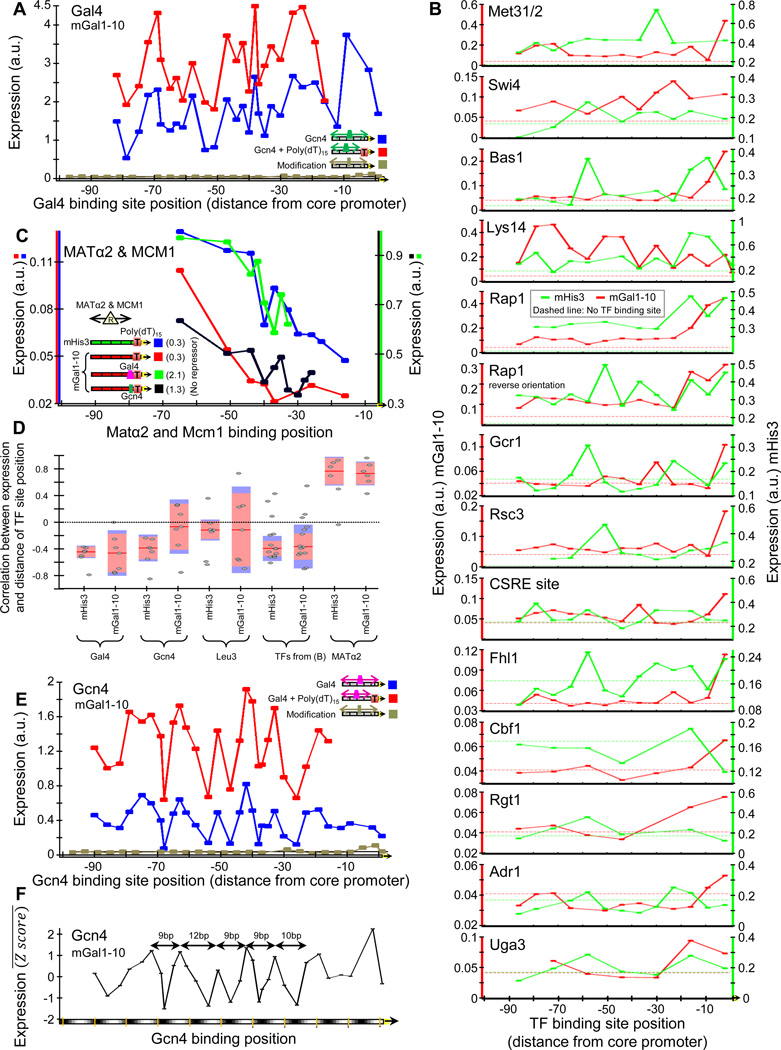
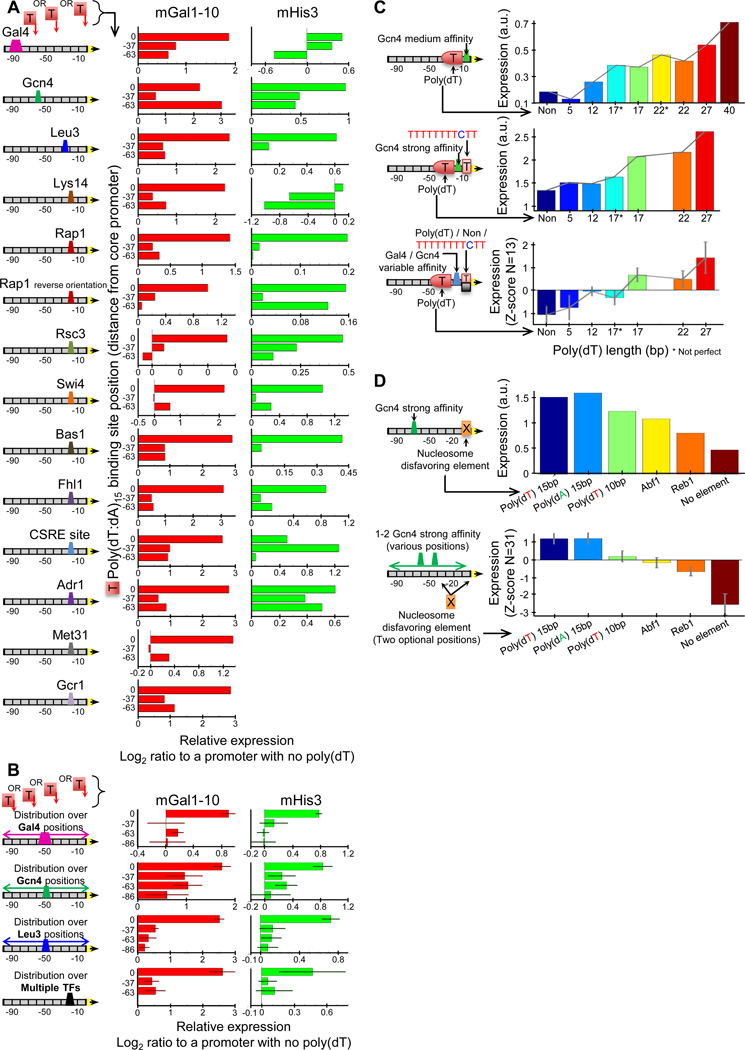
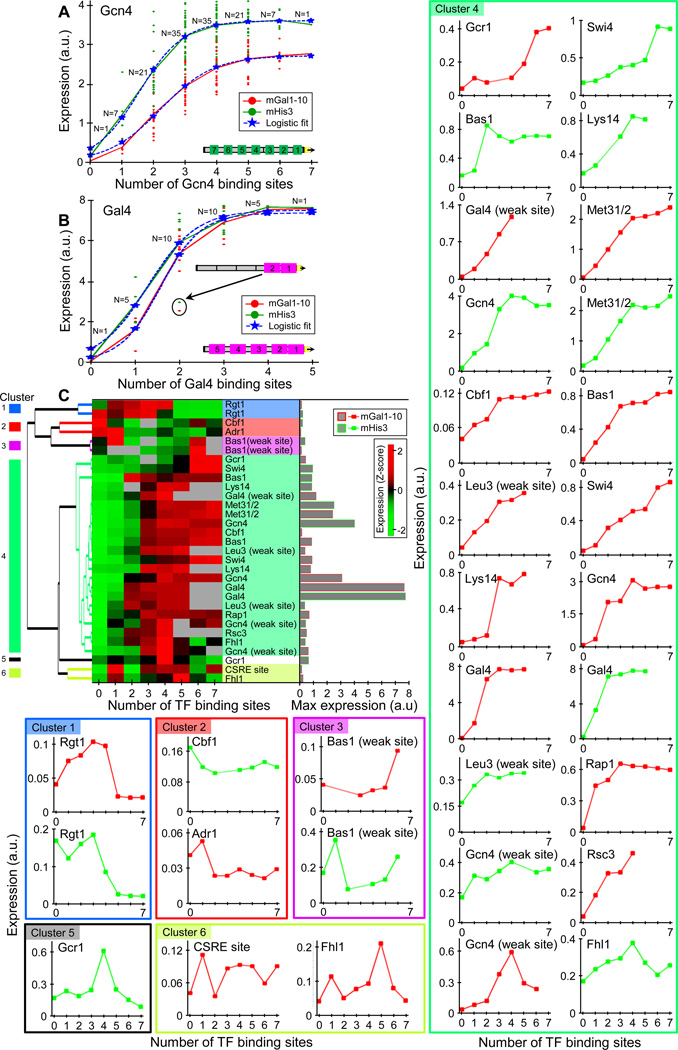
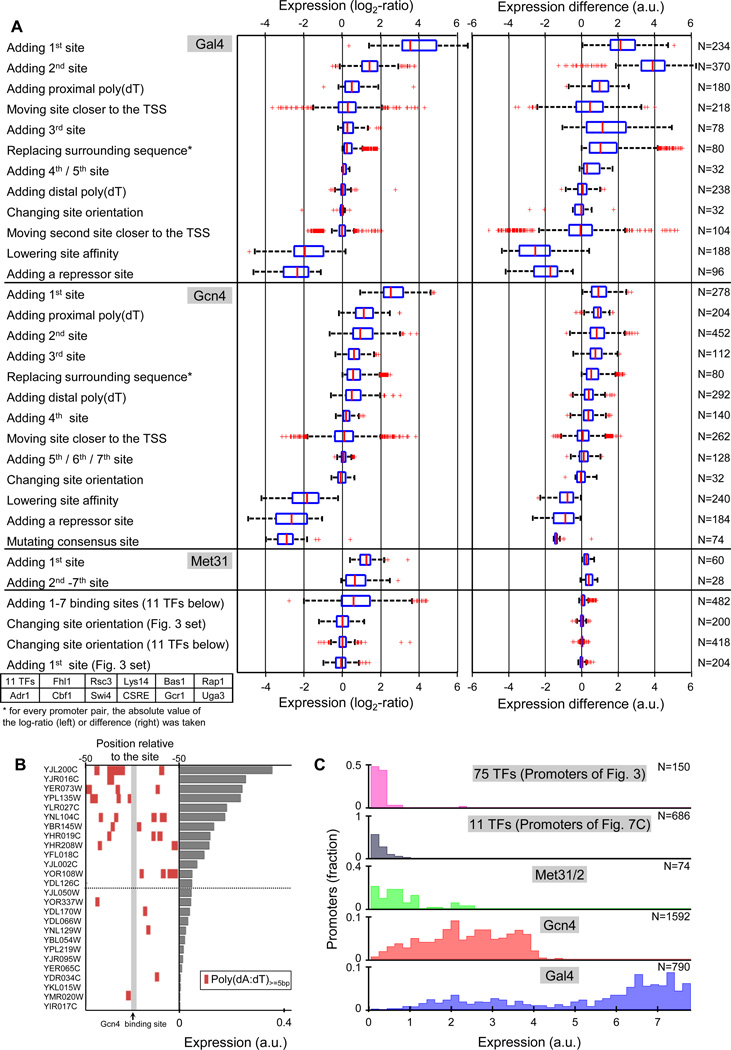
Despite extensive research, our understanding of the rules according to which cis-regulatory sequences are converted into gene expression is limited. We devised a method for obtaining parallel, highly accurate gene expression measurements from thousands of designed promoters and applied it to measure the effect of systematic changes in the location, number, orientation, affinity and organization of transcription-factor binding sites and nucleosome-disfavoring sequences. Our analyses reveal a clear relationship between expression and binding-site multiplicity, as well as dependencies of expression on the distance between transcription-factor binding sites and gene starts which are transcription-factor specific, including a striking ∼10-bp periodic relationship between gene expression and binding-site location. We show how this approach can measure transcription-factor sequence specificities and the sensitivity of transcription-factor sites to the surrounding sequence context, and compare the activity of 75 yeast transcription factors. Our method can be used to study both cis and trans effects of genotype on transcriptional, post-transcriptional and translational control.
Figures
Comment in
-
Dissecting genomic regulatory elements in vivo.Nat Biotechnol. 2012 Jun 7;30(6):504-6. doi: 10.1038/nbt.2266. Nat Biotechnol. 2012. PMID: 22678387 No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
