

Mammalian iron metabolism and its control by iron regulatory proteins
- PMID: 22610083
- PMCID: PMC3675657
- DOI: 10.1016/j.bbamcr.2012.05.010
Mammalian iron metabolism and its control by iron regulatory proteins
Abstract
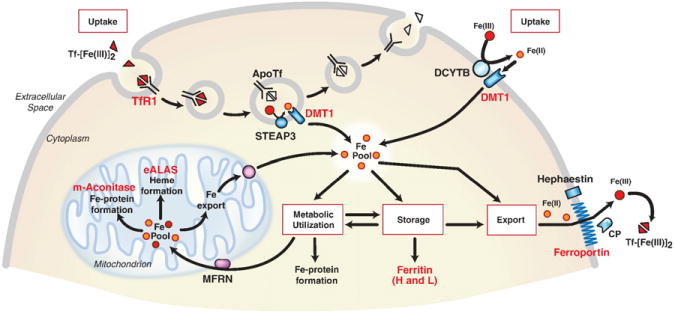
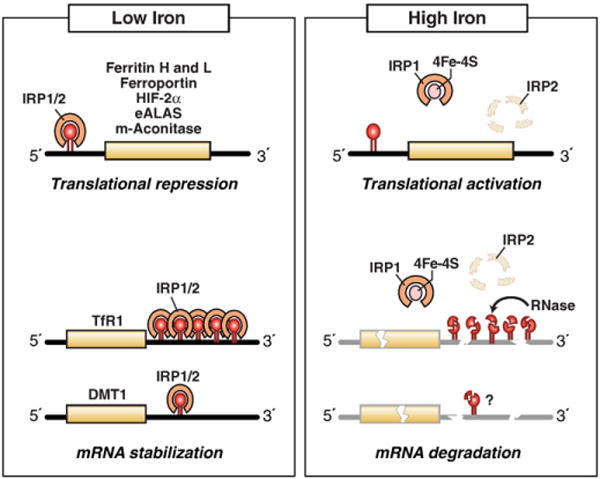
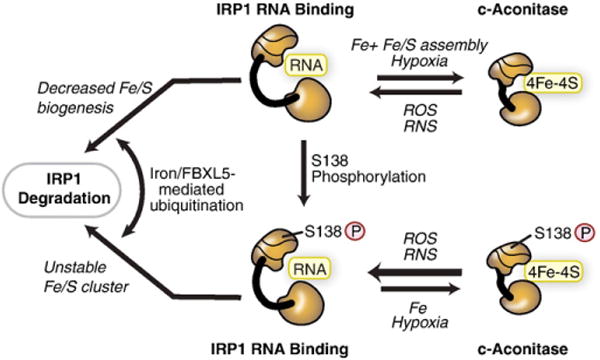
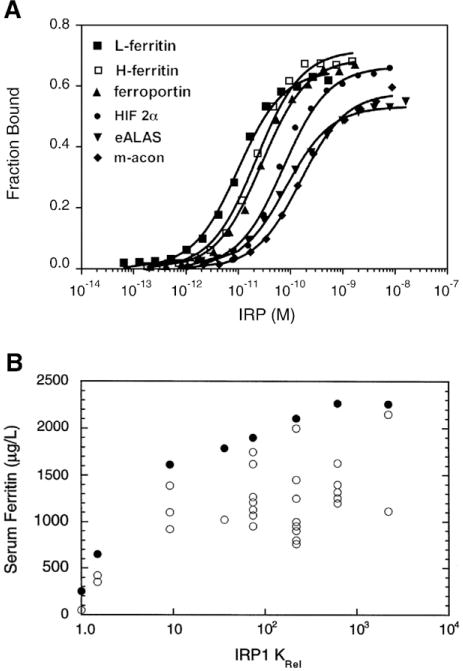
Cellular iron homeostasis is maintained by iron regulatory proteins 1 and 2 (IRP1 and IRP2). IRPs bind to iron-responsive elements (IREs) located in the untranslated regions of mRNAs encoding protein involved in iron uptake, storage, utilization and export. Over the past decade, significant progress has been made in understanding how IRPs are regulated by iron-dependent and iron-independent mechanisms and the pathological consequences of IRP2 deficiency in mice. The identification of novel IREs involved in diverse cellular pathways has revealed that the IRP-IRE network extends to processes other than iron homeostasis. A mechanistic understanding of IRP regulation will likely yield important insights into the basis of disorders of iron metabolism. This article is part of a Special Issue entitled: Cell Biology of Metals.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures
References
-
- Fleming RE, Ponka P. Iron overload in human disease. N Engl J Med. 2012;366:348–359. - PubMed
-
- Madsen E, Gitlin JD. Copper and iron disorders of the brain. Annu Rev Neurosci. 2007;30:317–337. - PubMed
-
- Simmons DA, Casale M, Alcon B, Pham N, Narayan N, Lynch G. Ferritin accumulation in dystrophic microglia is an early event in the development of Huntington’s disease. Glia. 2007;55:1074–1084. - PubMed
-
- Rouault TA. The role of iron regulatory proteins in mammalian iron homeostasis and disease. Nat Chem Biol. 2006;2:406–414. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 GM045201/GM/NIGMS NIH HHS/United States
- DK068602/DK/NIDDK NIH HHS/United States
- R01 DK068602/DK/NIDDK NIH HHS/United States
- T32DK007115/DK/NIDDK NIH HHS/United States
- R56 DK089212/DK/NIDDK NIH HHS/United States
- GM45201/GM/NIGMS NIH HHS/United States
- T32 DK007665/DK/NIDDK NIH HHS/United States
- DK089212/DK/NIDDK NIH HHS/United States
- R01 DK066600/DK/NIDDK NIH HHS/United States
- T32DK007665/DK/NIDDK NIH HHS/United States
- R56 DK066600/DK/NIDDK NIH HHS/United States
- T32 DK007115/DK/NIDDK NIH HHS/United States
- DK66600/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
