

S-adenosylmethionine decarboxylase overexpression inhibits mouse skin tumor promotion
- PMID: 22610166
- PMCID: PMC3499050
- DOI: 10.1093/carcin/bgs184
S-adenosylmethionine decarboxylase overexpression inhibits mouse skin tumor promotion
Abstract
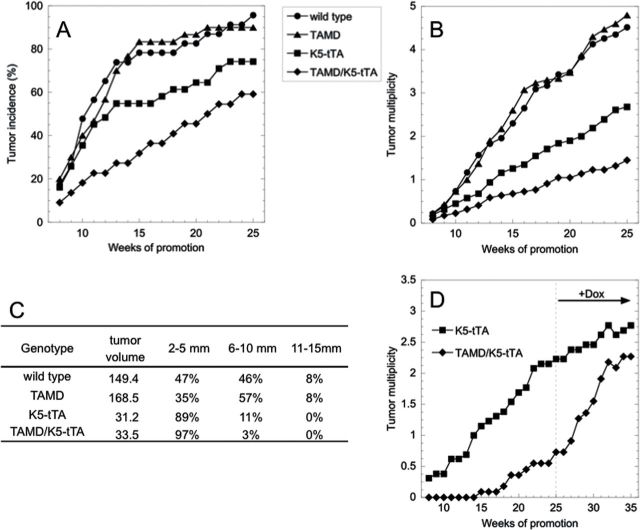
Neoplastic growth is associated with increased polyamine biosynthetic activity and content. Tumor promoter treatment induces the rate-limiting enzymes in polyamine biosynthesis, ornithine decarboxylase (ODC), and S-adenosylmethionine decarboxylase (AdoMetDC), and targeted ODC overexpression is sufficient for tumor promotion in initiated mouse skin. We generated a mouse model with doxycycline (Dox)-regulated AdoMetDC expression to determine the impact of this second rate-limiting enzyme on epithelial carcinogenesis. TetO-AdoMetDC (TAMD) transgenic founders were crossed with transgenic mice (K5-tTA) that express the tetracycline-regulated transcriptional activator within basal keratinocytes of the skin. Transgene expression in TAMD/K5-tTA mice was restricted to keratin 5 (K5) target tissues and silenced upon Dox treatment. AdoMetDC activity and its product, decarboxylated AdoMet, both increased approximately 8-fold in the skin. This enabled a redistribution of the polyamines that led to reduced putrescine, increased spermine, and an elevated spermine:spermidine ratio. Given the positive association between polyamine biosynthetic capacity and neoplastic growth, it was somewhat surprising to find that TAMD/K5-tTA mice developed significantly fewer tumors than controls in response to 7,12-dimethylbenz[a]anthracene/12-O-tetradecanoylphorbol-13-acetate chemical carcinogenesis. Importantly, tumor counts in TAMD/K5-tTA mice rebounded to nearly equal the levels in the control group upon Dox-mediated transgene silencing at a late stage of tumor promotion, which indicates that latent viable initiated cells remain in AdoMetDC-expressing skin. These results underscore the complexity of polyamine modulation of tumor development and emphasize the critical role of putrescine in tumor promotion. AdoMetDC-expressing mice will enable more refined spatial and temporal manipulation of polyamine biosynthesis during tumorigenesis and in other models of human disease.
Figures


Similar articles
-
Spermine synthase overexpression in vivo does not increase susceptibility to DMBA/TPA skin carcinogenesis or Min-Apc intestinal tumorigenesis.Cancer Biol Ther. 2012 Apr;13(6):358-68. doi: 10.4161/cbt.19241. Epub 2012 Apr 1. Cancer Biol Ther. 2012. PMID: 22258329 Free PMC article.
-
Targeted antizyme expression in the skin of transgenic mice reduces tumor promoter induction of ornithine decarboxylase and decreases sensitivity to chemical carcinogenesis.Cancer Res. 2001 Aug 15;61(16):6073-81. Cancer Res. 2001. PMID: 11507056
-
Effects of S-adenosyl-1,8-diamino-3-thio-octane and S-methyl-5'-methylthioadenosine on polyamine synthesis in Ehrlich ascites-tumour cells.Biochem J. 1989 Jul 1;261(1):205-10. doi: 10.1042/bj2610205. Biochem J. 1989. PMID: 2775206 Free PMC article.
-
Translational regulation of ornithine decarboxylase and other enzymes of the polyamine pathway.Int J Biochem Cell Biol. 1999 Jan;31(1):107-22. doi: 10.1016/s1357-2725(98)00135-6. Int J Biochem Cell Biol. 1999. PMID: 10216947 Review.
-
Polyamine metabolism.Digestion. 1990;46 Suppl 2:319-30. doi: 10.1159/000200405. Digestion. 1990. PMID: 2262065 Review.
Cited by
-
Elevated ornithine decarboxylase activity promotes skin tumorigenesis by stimulating the recruitment of bulge stem cells but not via toxic polyamine catabolic metabolites.Amino Acids. 2014 Mar;46(3):543-52. doi: 10.1007/s00726-013-1559-0. Epub 2013 Jul 25. Amino Acids. 2014. PMID: 23884694 Free PMC article.
-
Polyamines and cancer: implications for chemotherapy and chemoprevention.Expert Rev Mol Med. 2013 Feb 22;15:e3. doi: 10.1017/erm.2013.3. Expert Rev Mol Med. 2013. PMID: 23432971 Free PMC article. Review.
-
A Simple and Efficient Method for the Substrate Identification of Amino Acid Decarboxylases.Int J Mol Sci. 2022 Nov 22;23(23):14551. doi: 10.3390/ijms232314551. Int J Mol Sci. 2022. PMID: 36498879 Free PMC article.
-
Expression and distribution patterns of spermine, spermidine, and putrescine in rat hair follicle.Histochem Cell Biol. 2018 Feb;149(2):161-167. doi: 10.1007/s00418-017-1621-1. Epub 2017 Nov 21. Histochem Cell Biol. 2018. PMID: 29159700
-
Skin Carcinogenesis Studies Using Mouse Models with Altered Polyamines.Cancer Growth Metastasis. 2015 Aug 9;8(Suppl 1):17-27. doi: 10.4137/CGM.S21219. eCollection 2015. Cancer Growth Metastasis. 2015. PMID: 26380554 Free PMC article. Review.
References
-
- Igarashi K., et al. Modulation of cellular function by polyamines. Int. J. Biochem. Cell Biol. (2010);42:39–51. - PubMed
-
- Basuroy U.K., et al. Emerging concepts in targeting the polyamine metabolic pathway in epithelial cancer chemoprevention and chemotherapy. J. Biochem. (2006);139:27–33. - PubMed
-
- Casero R.A., Jr, et al. Targeting polyamine metabolism and function in cancer and other hyperproliferative diseases. Nat. Rev. Drug Discov. (2007);6:373–390. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
