

Nascent RNA structure modulates the transcriptional dynamics of RNA polymerases
- PMID: 22615360
- PMCID: PMC3384149
- DOI: 10.1073/pnas.1205063109
Nascent RNA structure modulates the transcriptional dynamics of RNA polymerases
Abstract
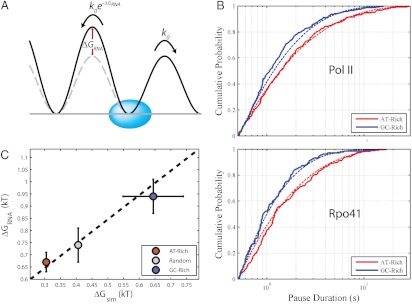
RNA polymerase pausing represents an important mechanism of transcriptional regulation. In this study, we use a single-molecule transcription assay to investigate the effect of template base-pair composition on pausing by RNA polymerase II and the evolutionarily distinct mitochondrial polymerase Rpo41. For both enzymes, pauses are shorter and less frequent on GC-rich templates. Significantly, incubation with RNase abolishes the template dependence of pausing. A kinetic model, wherein the secondary structure of the nascent RNA poses an energetic barrier to pausing by impeding backtracking along the template, quantitatively predicts the pause densities and durations observed. The energy barriers extracted from the data correlate well with RNA folding energies obtained from cotranscriptional folding simulations. These results reveal that RNA secondary structures provide a cis-acting mechanism by which sequence modulates transcriptional elongation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Rpb5 modulates the RNA polymerase II transition from initiation to elongation by influencing Spt5 association and backtracking.Biochim Biophys Acta Gene Regul Mech. 2018 Jan;1861(1):1-13. doi: 10.1016/j.bbagrm.2017.11.002. Epub 2017 Nov 11. Biochim Biophys Acta Gene Regul Mech. 2018. PMID: 29133017
-
Yeast DEAD box protein Mss116p is a transcription elongation factor that modulates the activity of mitochondrial RNA polymerase.Mol Cell Biol. 2014 Jul;34(13):2360-9. doi: 10.1128/MCB.00160-14. Epub 2014 Apr 14. Mol Cell Biol. 2014. PMID: 24732805 Free PMC article.
-
Intrinsic translocation barrier as an initial step in pausing by RNA polymerase II.J Mol Biol. 2013 Feb 22;425(4):697-712. doi: 10.1016/j.jmb.2012.12.002. Epub 2012 Dec 10. J Mol Biol. 2013. PMID: 23238253 Free PMC article.
-
Transcriptional Pausing as a Mediator of Bacterial Gene Regulation.Annu Rev Microbiol. 2021 Oct 8;75:291-314. doi: 10.1146/annurev-micro-051721-043826. Epub 2021 Aug 4. Annu Rev Microbiol. 2021. PMID: 34348029 Review.
-
Pause & go: from the discovery of RNA polymerase pausing to its functional implications.Curr Opin Cell Biol. 2017 Jun;46:72-80. doi: 10.1016/j.ceb.2017.03.002. Epub 2017 Mar 28. Curr Opin Cell Biol. 2017. PMID: 28363125 Free PMC article. Review.
Cited by
-
TEFM Enhances Transcription Elongation by Modifying mtRNAP Pausing Dynamics.Biophys J. 2018 Dec 18;115(12):2295-2300. doi: 10.1016/j.bpj.2018.11.004. Epub 2018 Nov 10. Biophys J. 2018. PMID: 30514634 Free PMC article.
-
Stochastic approach to the molecular counting problem in superresolution microscopy.Proc Natl Acad Sci U S A. 2015 Jan 13;112(2):E110-8. doi: 10.1073/pnas.1408071112. Epub 2014 Dec 22. Proc Natl Acad Sci U S A. 2015. PMID: 25535361 Free PMC article.
-
Single molecule microscopy reveals mechanistic insight into RNA polymerase II preinitiation complex assembly and transcriptional activity.Nucleic Acids Res. 2016 Sep 6;44(15):7132-43. doi: 10.1093/nar/gkw321. Epub 2016 Apr 25. Nucleic Acids Res. 2016. PMID: 27112574 Free PMC article.
-
Sequence-specific thermodynamic properties of nucleic acids influence both transcriptional pausing and backtracking in yeast.PLoS One. 2017 Mar 16;12(3):e0174066. doi: 10.1371/journal.pone.0174066. eCollection 2017. PLoS One. 2017. PMID: 28301878 Free PMC article.
-
Precursor RNA structural patterns at SF3B1 mutation sensitive cryptic 3' splice sites.bioRxiv [Preprint]. 2025 Feb 22:2025.02.19.638873. doi: 10.1101/2025.02.19.638873. bioRxiv. 2025. PMID: 40027643 Free PMC article. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
