

Intron-mediated alternative splicing of Arabidopsis P5CS1 and its association with natural variation in proline and climate adaptation
- PMID: 22615385
- PMCID: PMC3384178
- DOI: 10.1073/pnas.1203433109
Intron-mediated alternative splicing of Arabidopsis P5CS1 and its association with natural variation in proline and climate adaptation
Abstract
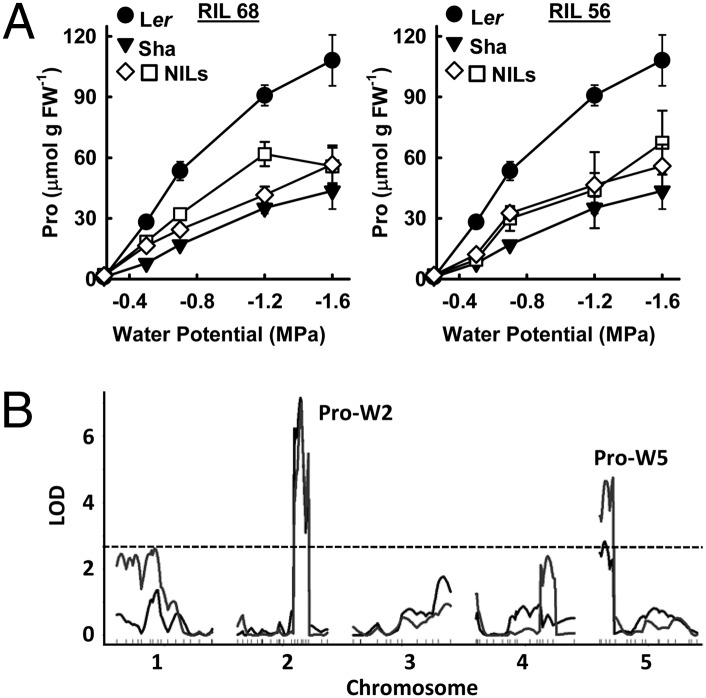
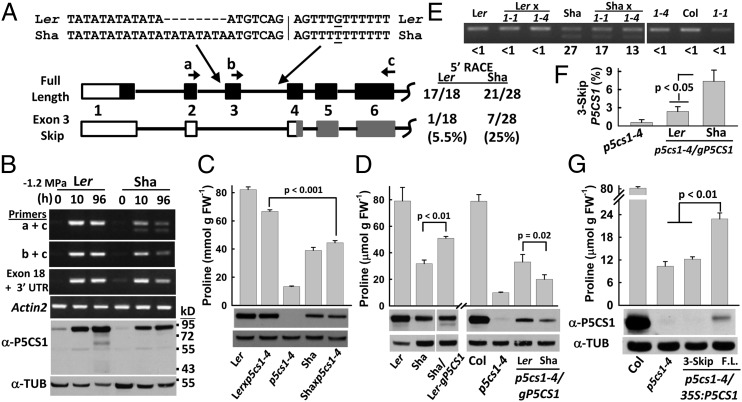
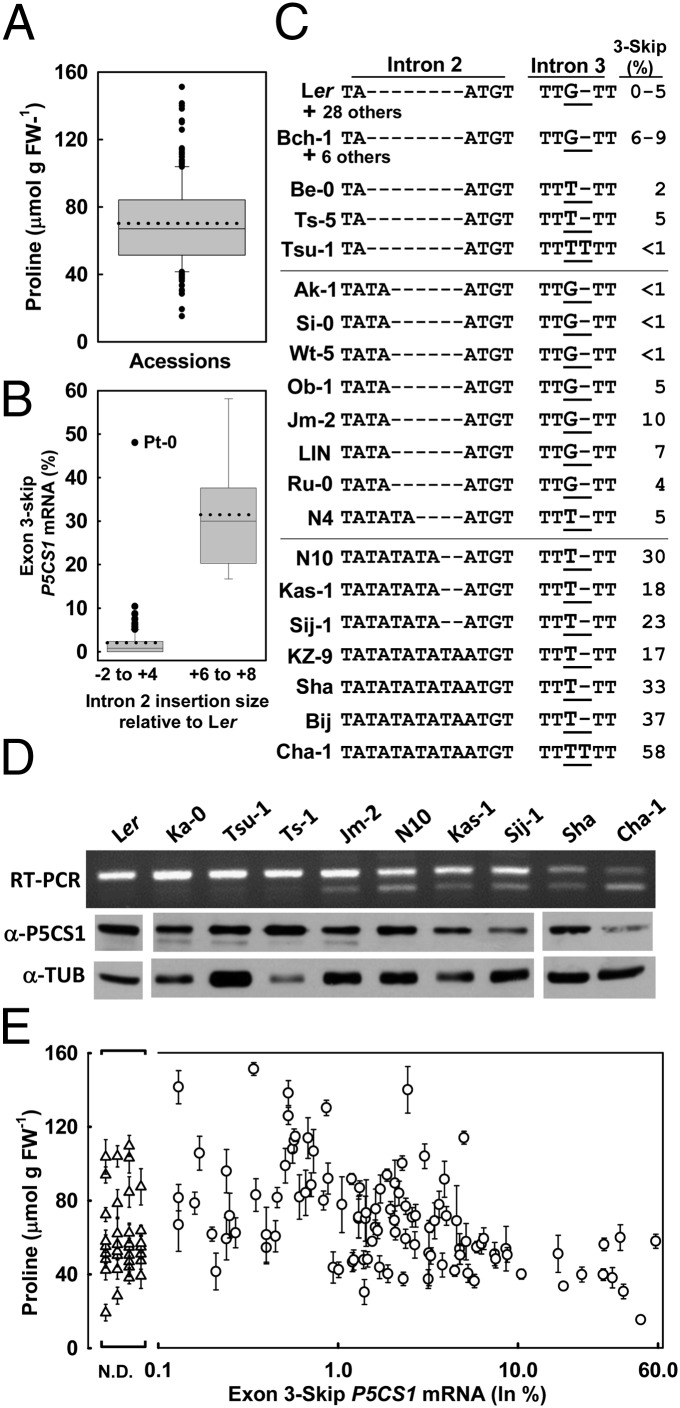
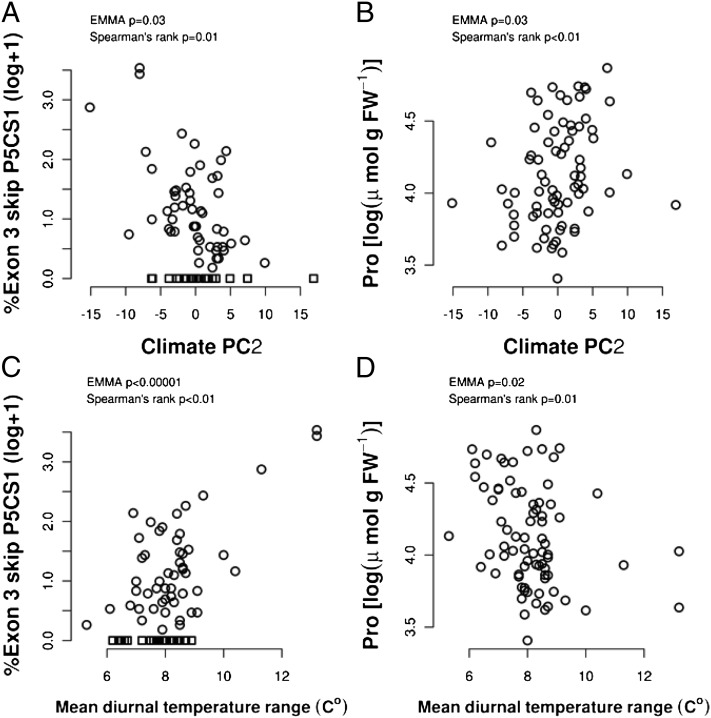
Drought-induced proline accumulation is widely observed in plants but its regulation and adaptive value are not as well understood. Proline accumulation of the Arabidopsis accession Shakdara (Sha) was threefold less than that of Landsberg erecta (Ler) and quantitative trait loci mapping identified a reduced function allele of the proline synthesis enzyme Δ(1)-pyrroline-5-carboxylate synthetase1 (P5CS1) as a basis for the lower proline of Sha. Sha P5CS1 had additional TA repeats in intron 2 and a G-to-T transversion in intron 3 that were sufficient to promote alternative splicing and production of a nonfunctional transcript lacking exon 3 (exon 3-skip P5CS1). In Sha, and additional accessions with the same intron polymorphisms, the nonfunctional exon 3-skip P5CS1 splice variant constituted as much as half of the total P5CS1 transcript. In a larger panel of Arabidopsis accessions, low water potential-induced proline accumulation varied by 10-fold and variable production of exon 3-skip P5CS1 among accessions was an important, but not the sole, factor underlying variation in proline accumulation. Population genetic analyses suggest that P5CS1 may have evolved under positive selection, and more extensive correlation of exon 3-skip P5CS1 production than proline abundance with climate conditions of natural accessions also suggest a role of P5CS1 in local adaptation to the environment. These data identify a unique source of alternative splicing in plants, demonstrate a role of exon 3-skip P5CS1 in natural variation of proline metabolism, and suggest an association of P5CS1 and its alternative splicing with environmental adaptation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Szabados L, Savouré A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010;15:89–97. - PubMed
-
- Sharma S, Verslues PE. Mechanisms independent of abscisic acid (ABA) or proline feedback have a predominant role in transcriptional regulation of proline metabolism during low water potential and stress recovery. Plant Cell Environ. 2010;33:1838–1851. - PubMed
-
- Székely G, et al. Duplicated P5CS genes of Arabidopsis play distinct roles in stress regulation and developmental control of proline biosynthesis. Plant J. 2008;53:11–28. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
