

Requirement of neuronal connexin36 in pathways mediating presynaptic inhibition of primary afferents in functionally mature mouse spinal cord
- PMID: 22615430
- PMCID: PMC3476635
- DOI: 10.1113/jphysiol.2011.225987
Requirement of neuronal connexin36 in pathways mediating presynaptic inhibition of primary afferents in functionally mature mouse spinal cord
Abstract
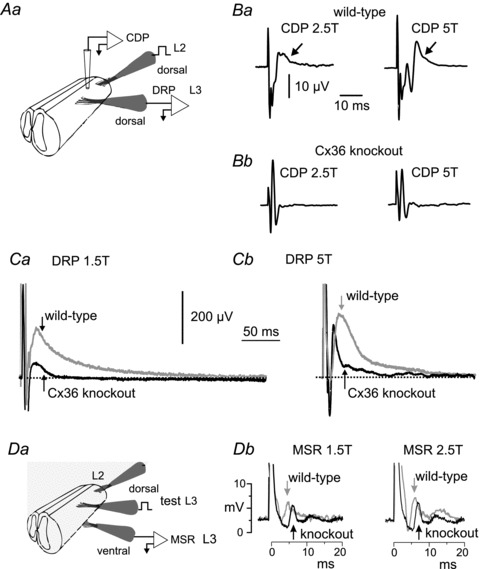
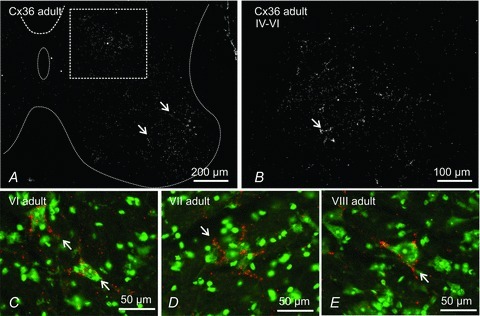
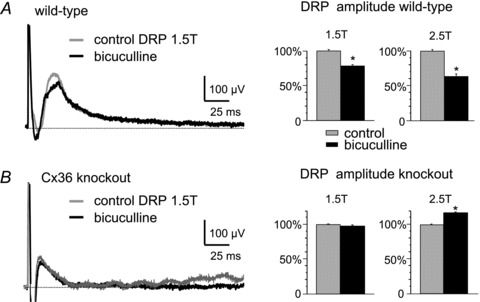
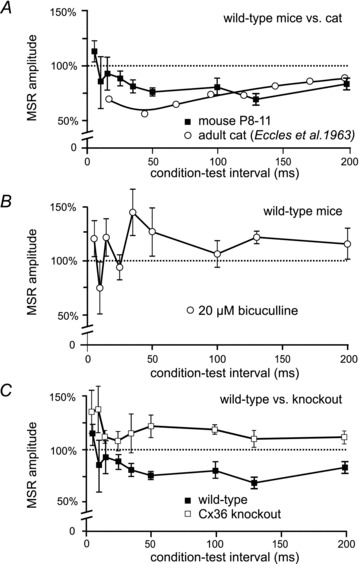
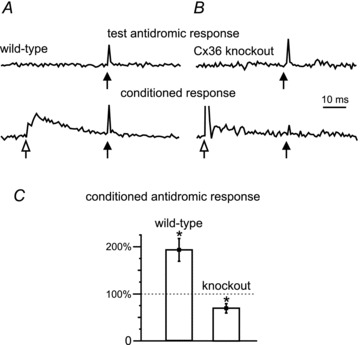
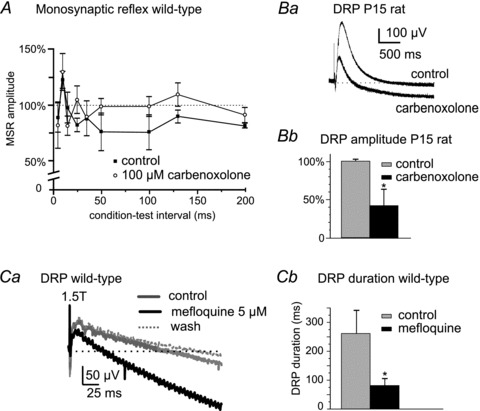
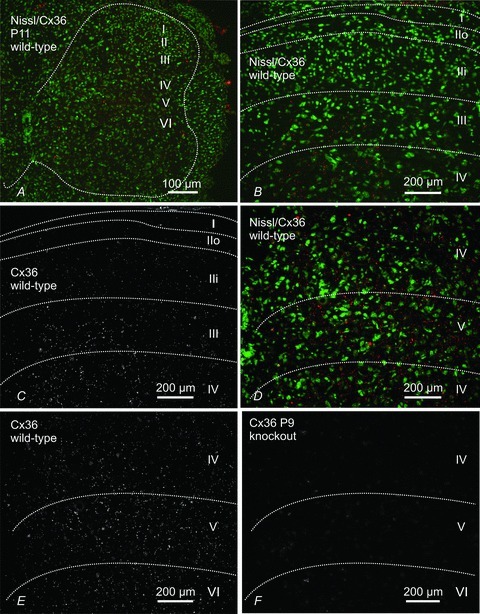
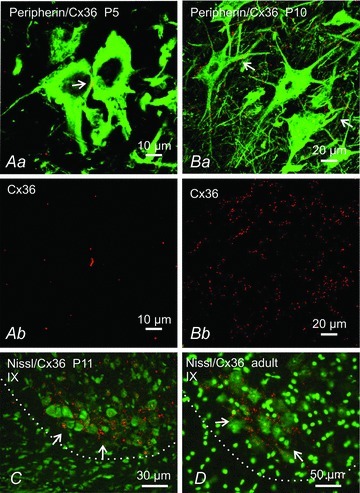
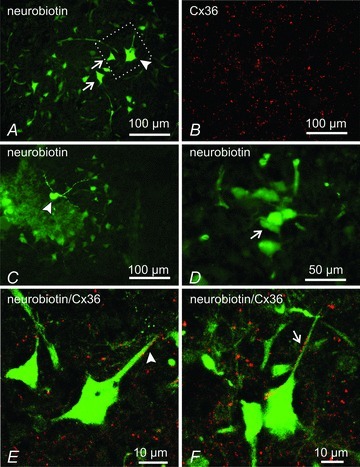
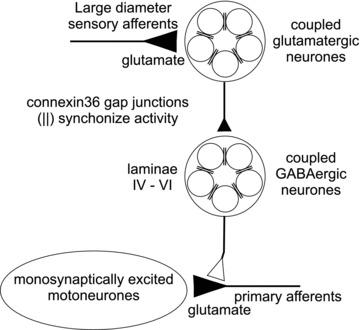
Electrical synapses formed by gap junctions containing connexin36 (Cx36) promote synchronous activity of interneurones in many regions of mammalian brain; however, there is limited information on the role of electrical synapses in spinal neuronal networks. Here we show that Cx36 is widely distributed in the spinal cord and is involved in mechanisms that govern presynaptic inhibition of primary afferent terminals. Electrophysiological recordings were made in spinal cord preparations from 8- to 11-day-old wild-type and Cx36 knockout mice. Several features associated with presynaptic inhibition evoked by conditioning stimulation of low threshold hindlimb afferents were substantially compromised in Cx36 knockout mice. Dorsal root potentials (DRPs) evoked by low intensity stimulation of sensory afferents were reduced in amplitude by 79% and in duration by 67% in Cx36 knockouts. DRPs were similarly affected in wild-types by bath application of gap junction blockers. Consistent with presynaptic inhibition of group Ia muscle spindle afferent terminals on motoneurones described in adult cats, conditioning stimulation of an adjacent dorsal root evoked a long duration inhibition of monosynaptic reflexes recorded from the ventral root in wild-type mice, and this inhibition was antagonized by bicuculline. The same conditioning stimulation failed to inhibit monosynaptic reflexes in Cx36 knockout mice. Immunofluorescence labelling for Cx36 was found throughout the dorsal and ventral horns of the spinal cord of juvenile mice and persisted in mature animals. In deep dorsal horn laminae, where interneurones involved in presynaptic inhibition of large diameter muscle afferents are located, cells were extensively dye-coupled following intracellular neurobiotin injection. Coupled cells displayed Cx36-positive puncta along their processes. Our results indicate that gap junctions formed by Cx36 in spinal cord are required for maintenance of presynaptic inhibition, including the regulation of transmission from Ia muscle spindle afferents. In addition to a role in presynaptic inhibition in juvenile animals, the persistence of Cx36 expression among spinal neuronal populations in the adult mouse suggests that the contribution of electrical synapses to integrative processes in fully mature spinal cord may be as diverse as that found in other areas of the CNS.
Figures
References
-
- Bagust J, Green KA, Kerkut GA. Strychnine sensitive inhibition in the dorsal horn of mammalian spinal cord. Brain Res. 1981;217:425–429. - PubMed
-
- Bagust J, Forsythe D, Kerkut GA. Demonstration of the synaptic origin of primary afferent depolarisation (PAD) in the isolated spinal cord of the hamster. Brain Res. 1985;341:385–389. - PubMed
-
- Bennett MVL. Gap junctions as electrical synapses. J Neurocytol. 1997;26:349–366. - PubMed
-
- Bennett MV, Zukin RS. Electrical coupling and neuronal synchronization in the mammalian brain. Neuron. 2004;41:495–511. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous