

Mechanisms and evolutionary patterns of mammalian and avian dosage compensation
- PMID: 22615540
- PMCID: PMC3352821
- DOI: 10.1371/journal.pbio.1001328
Mechanisms and evolutionary patterns of mammalian and avian dosage compensation
Abstract
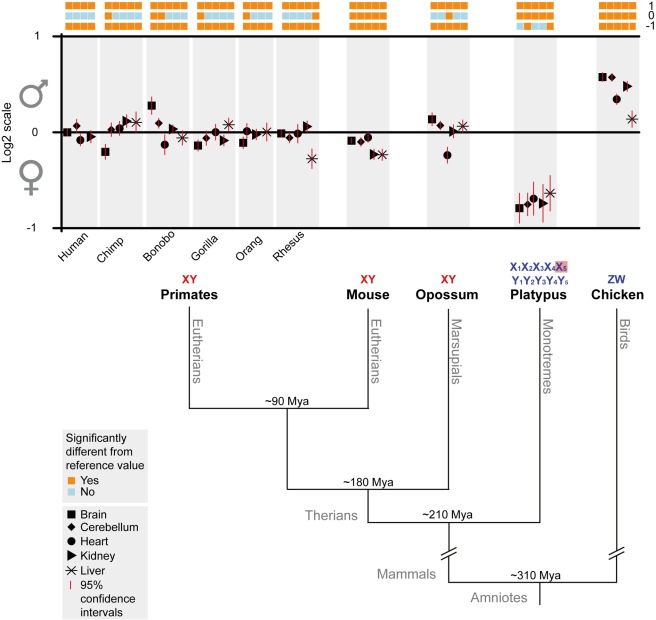
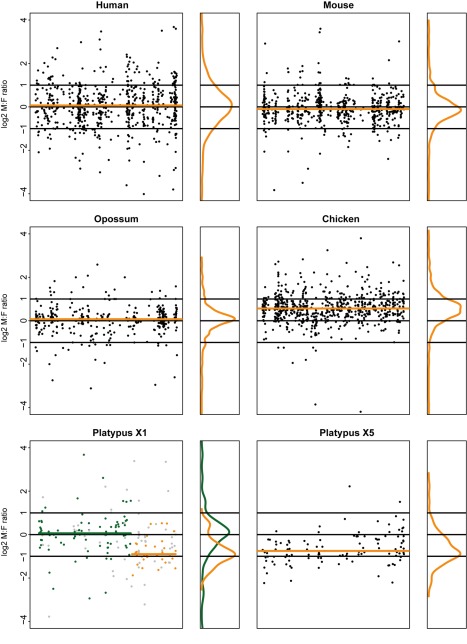
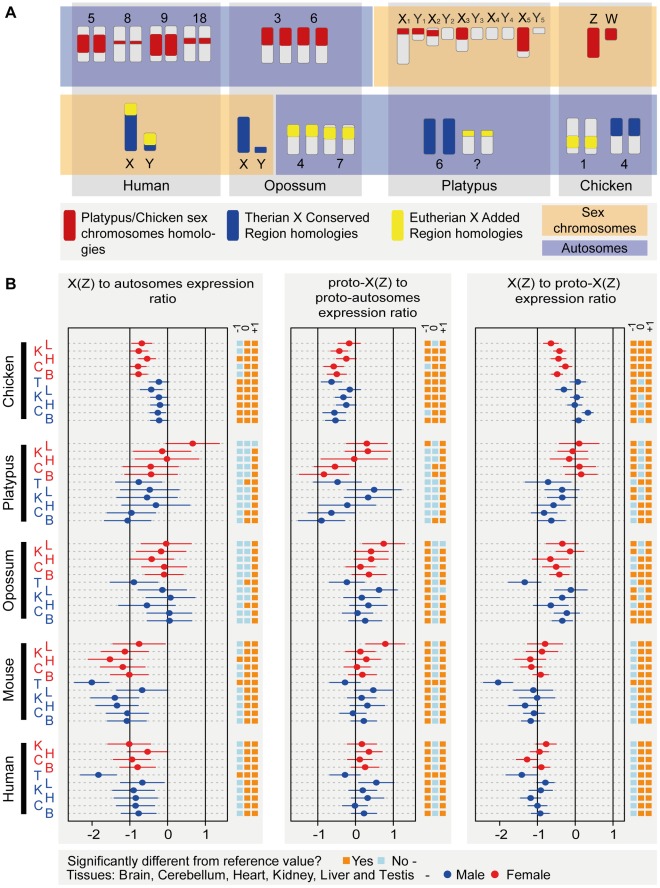
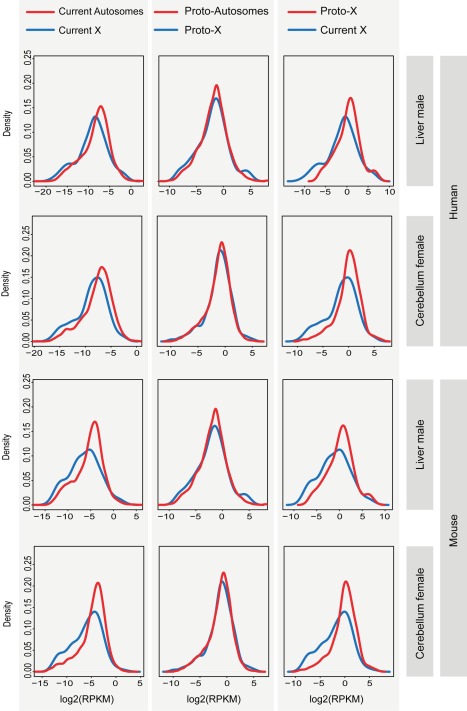
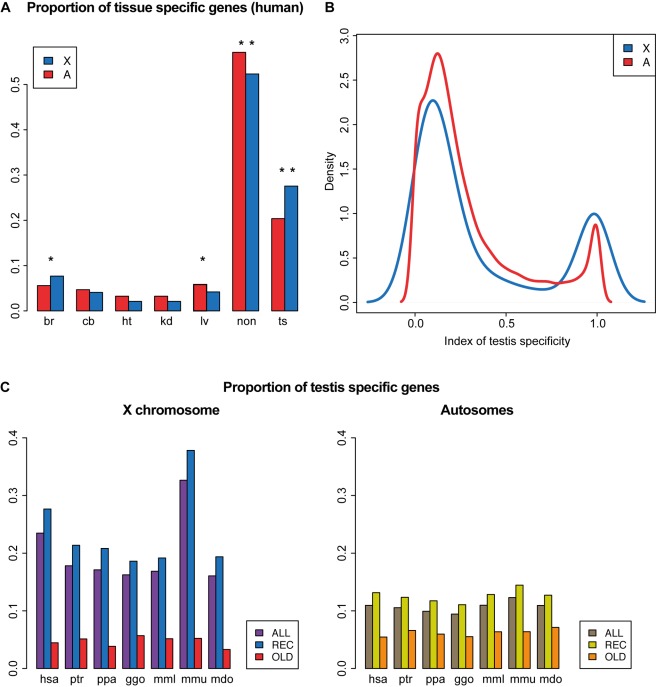
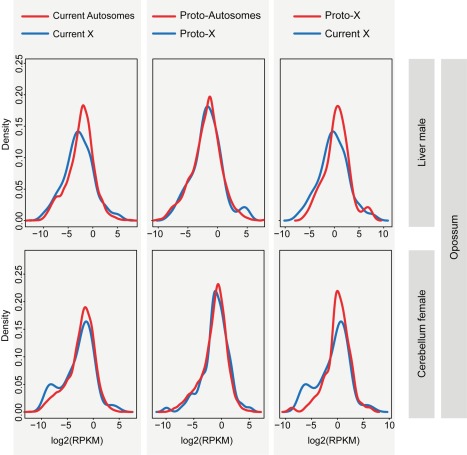
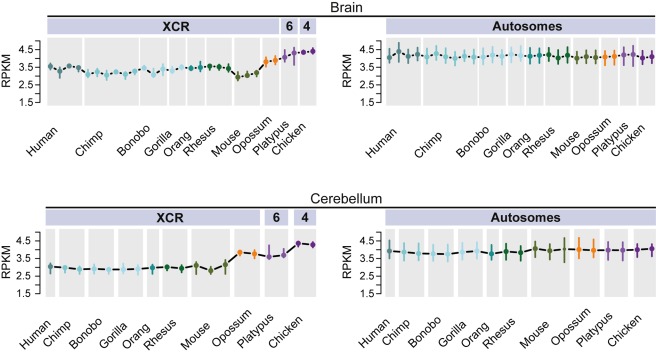
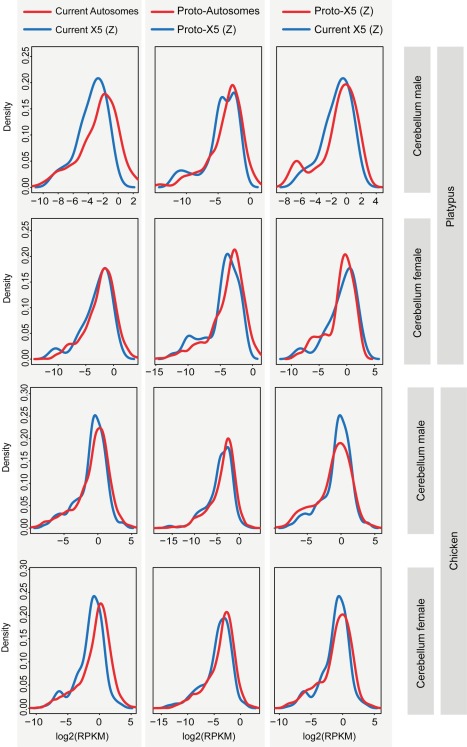
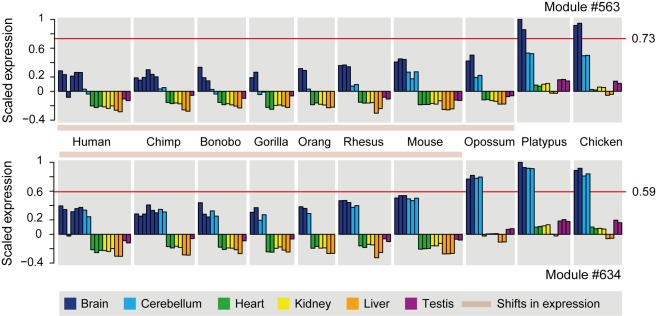
As a result of sex chromosome differentiation from ancestral autosomes, male mammalian cells only contain one X chromosome. It has long been hypothesized that X-linked gene expression levels have become doubled in males to restore the original transcriptional output, and that the resulting X overexpression in females then drove the evolution of X inactivation (XCI). However, this model has never been directly tested and patterns and mechanisms of dosage compensation across different mammals and birds generally remain little understood. Here we trace the evolution of dosage compensation using extensive transcriptome data from males and females representing all major mammalian lineages and birds. Our analyses suggest that the X has become globally upregulated in marsupials, whereas we do not detect a global upregulation of this chromosome in placental mammals. However, we find that a subset of autosomal genes interacting with X-linked genes have become downregulated in placentals upon the emergence of sex chromosomes. Thus, different driving forces may underlie the evolution of XCI and the highly efficient equilibration of X expression levels between the sexes observed for both of these lineages. In the egg-laying monotremes and birds, which have partially homologous sex chromosome systems, partial upregulation of the X (Z in birds) evolved but is largely restricted to the heterogametic sex, which provides an explanation for the partially sex-biased X (Z) expression and lack of global inactivation mechanisms in these lineages. Our findings suggest that dosage reductions imposed by sex chromosome differentiation events in amniotes were resolved in strikingly different ways.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Ohno S. Sex chromosomes and sex-linked genes. Berlin: Springer-Verlag; 1967.
-
- Potrzebowski L, Vinckenbosch N, Marques A. C, Chalmel F, Jegou B, et al. Chromosomal gene movements reflect the recent origin and biology of therian sex chromosomes. PLoS Biol. 2008;6:e80. doi: 10.1371/journal.pbio.0060080. - DOI - PMC - PubMed
-
- Grutzner F, Rens W, Tsend-Ayush E, El-Mogharbel N, O'Brien P. C, et al. In the platypus a meiotic chain of ten sex chromosomes shares genes with the bird Z and mammal X chromosomes. Nature. 2004;432:913–917. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
