

Role of N-terminal myristylation in the structure and regulation of cAMP-dependent protein kinase
- PMID: 22617327
- PMCID: PMC3597442
- DOI: 10.1016/j.jmb.2012.05.021
Role of N-terminal myristylation in the structure and regulation of cAMP-dependent protein kinase
Abstract
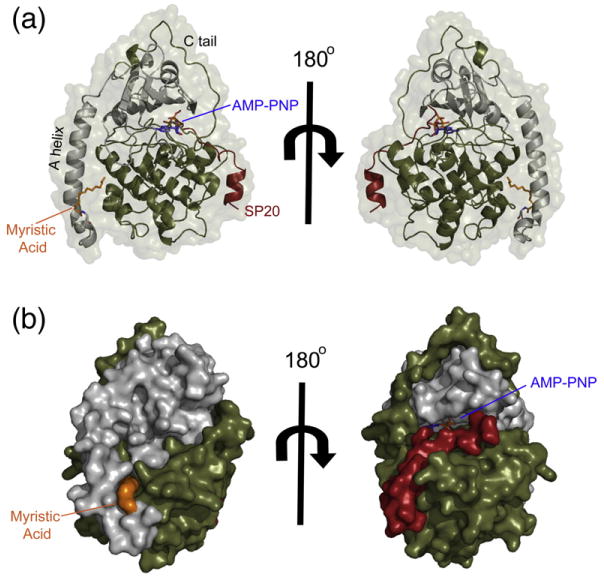
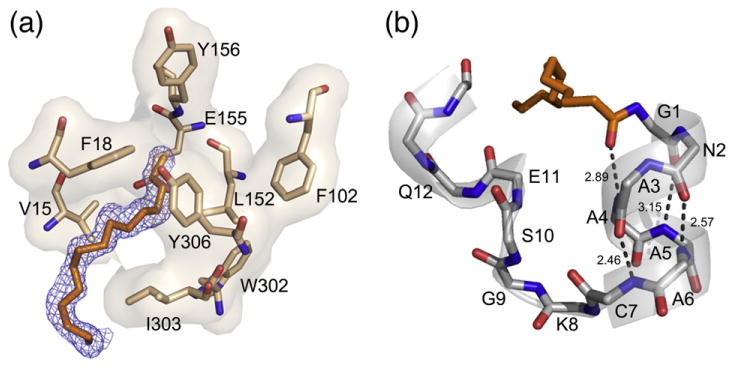
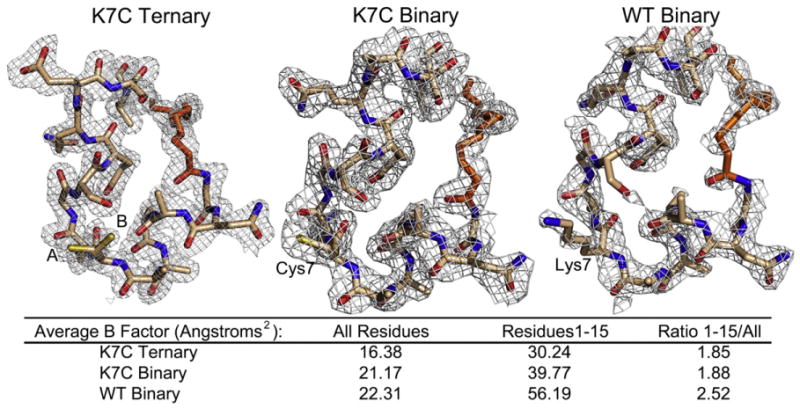
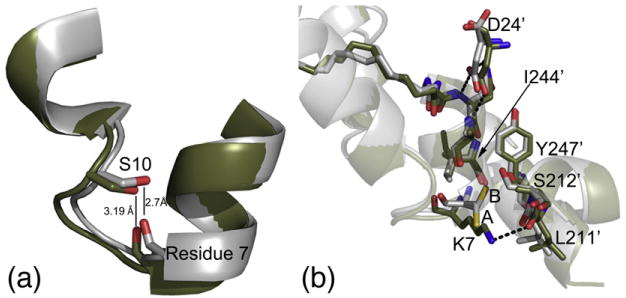
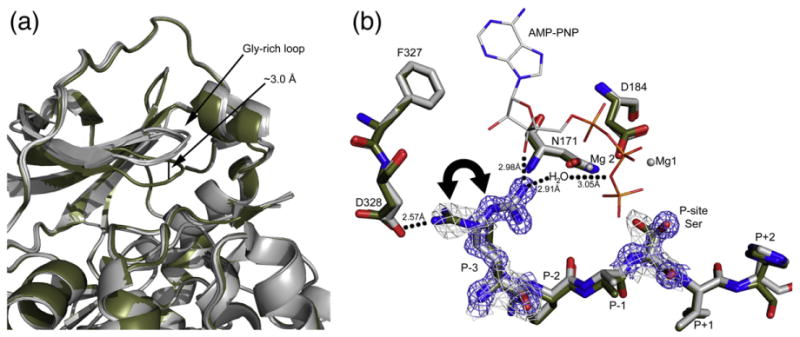
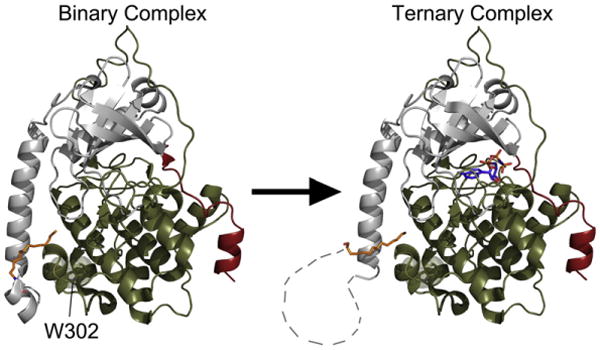
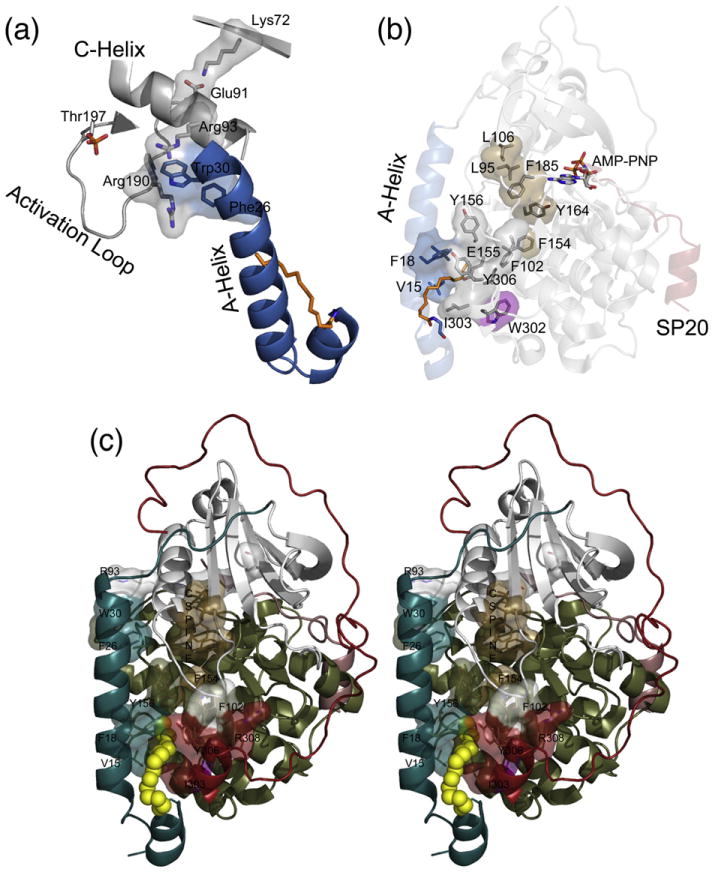
The catalytic (C) subunit of cAMP-dependent protein kinase [protein kinase A (PKA)] is a major target of cAMP signaling, and its regulation is of fundamental importance to biological processes. One mode of regulation is N-myristylation, which has eluded structural and functional characterization so far because most crystal structures are of the non-myristylated enzyme, are phosphorylated on Ser10, and generally lack electron density for the first 13 residues. We crystallized myristylated wild-type (WT) PKA and a K7C mutant as binary (bound to a substrate peptide) and ternary [bound to a substrate peptide and adenosine-5'-(β,γ-imido)triphosphate] complexes. There was clear electron density for the entire N-terminus in the binary complexes, both refined to 2.0 Å, and K7C ternary complex, refined to 1.35 Å. The N-termini in these three structures display a novel conformation with a previously unseen helix from residues 1 to 7. The K7C mutant appears to have a more stable N-terminus, and this correlated with a significant decrease in the B-factors for the N-terminus in the myr-K7C complexes compared to the WT binary complex. The N-terminus of the myristylated WT ternary complex, refined to 2.0 Å, was disordered as in previous structures. In addition to a more ordered N-terminus, the myristylated K7C mutant exhibited a 53% increase in k(cat). The effect of nucleotide binding on the structure of the N-terminus in the WT protein and the kinetic changes in the K7C protein suggest that myristylation or occupancy of the myristyl binding pocket may serve as a site for allosteric regulation in the C-subunit.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Figures
References
-
- Kirschner LS, Yin Z, Jones GN, Mahoney E. Mouse models of altered protein kinase A signaling. Endocr-Relat Cancer. 2009;16:773–793. - PubMed
-
- Mucignat-Caretta C, Caretta A. Clustered distribution of cAMP-dependent protein kinase regulatory isoform RI alpha during the development of the rat brain. J Comp Neurol. 2002;451:324–333. - PubMed
-
- Francis SH, Corbin JD. Structure and function of cyclic nucleotide-dependent protein kinases. Annu Rev Physiol. 1994;56:237–272. - PubMed
-
- Tasken K, Skalhegg BS, Tasken KA, Solberg R, Knutsen HK, Levy FO, et al. Structure, function, and regulation of human cAMP-dependent protein kinases. Adv Second Messenger Phosphoprotein Res. 1997;31:191–204. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
