

Computational model of a circulation current that controls electrochemical properties in the mammalian cochlea
- PMID: 22619324
- PMCID: PMC3384130
- DOI: 10.1073/pnas.1120067109
Computational model of a circulation current that controls electrochemical properties in the mammalian cochlea
Abstract
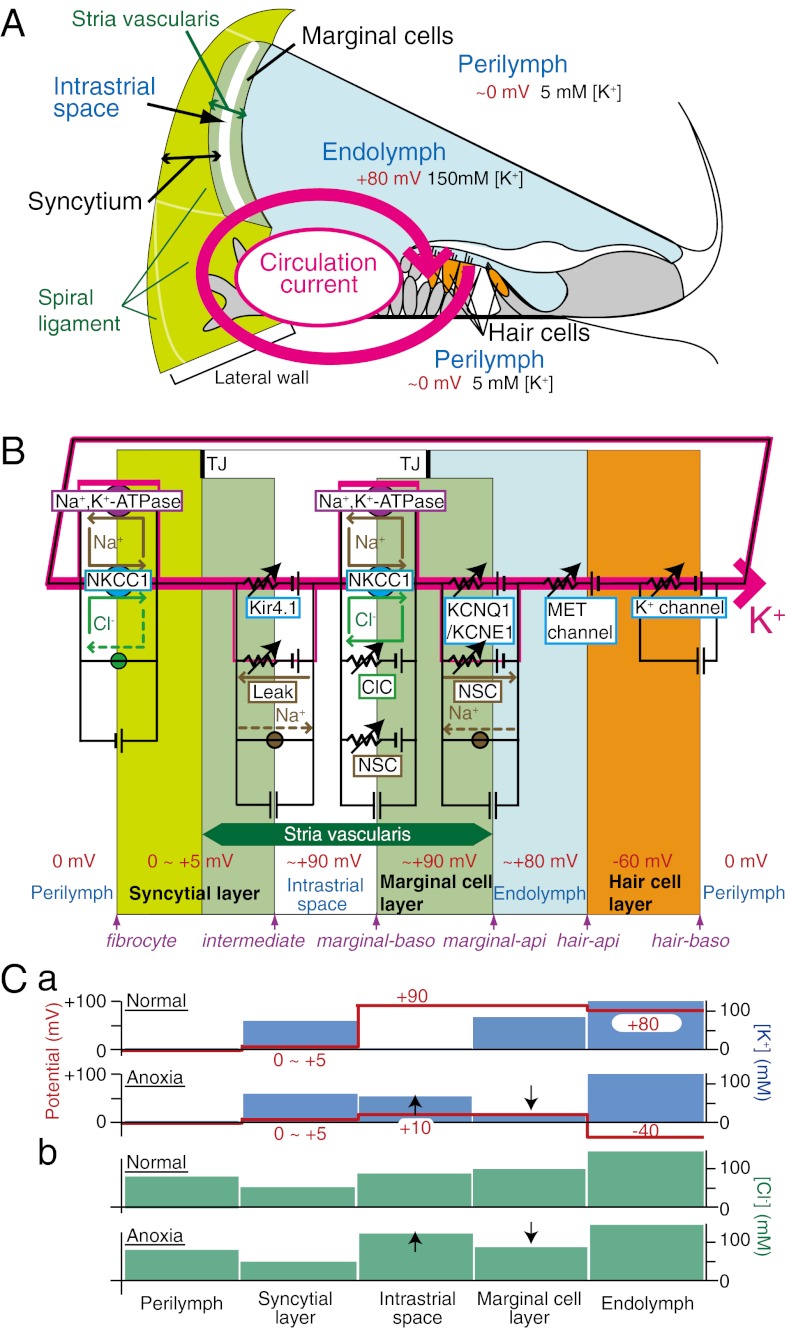
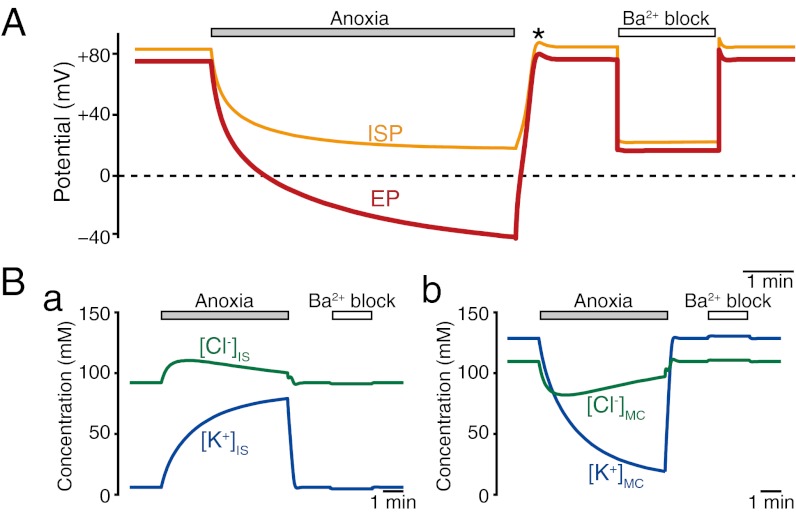
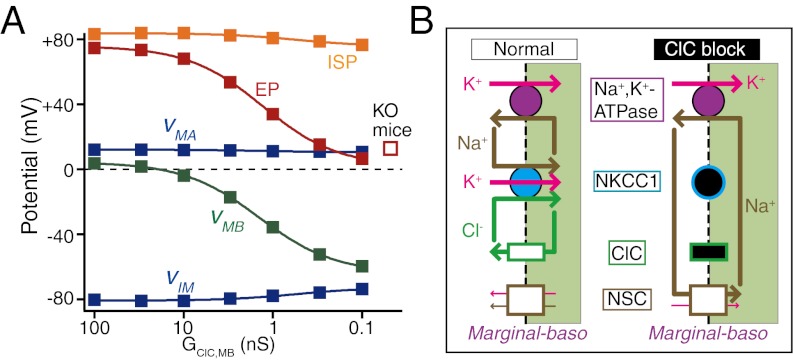
Sound-evoked mechanical stimuli permit endolymphatic K(+) to enter sensory hair cells. This transduction is sensitized by an endocochlear potential (EP) of +80 mV in endolymph. After depolarizing the cells, K(+) leaves hair cells in perilymph, and it is then circulated back to endolymph across the lateral cochlear wall. In theory, this process entails a continuous and unidirectional current carried by apical K(+) channels and basolateral K(+) uptake transporters in both the marginal cell and syncytial layers of the lateral wall. The transporters regulate intracellular and extracellular [K(+)], allowing the channels to form K(+) diffusion potentials across each of the two layers. These diffusion potentials govern the EP. What remains uncertain is whether these transport mechanisms accumulating across diverse cell layers make up a continuous circulation current in the lateral wall and how this current might affect the characteristics of the endolymph. To address this question, we developed an electrophysiological model that incorporates channels and transporters of the lateral wall and channels of hair cells that derive a circulation current. The simulation replicated normal experimental EP values and reproduced experimentally measured changes in the EP and intra- and extracellular [K(+)] in the lateral wall when different transporters and channels were blocked. The model predicts that, under these different conditions, the circulation current's contribution to the EP arises from different sources. Finally, our model also accurately simulated EP loss in a mouse model of a chloride channelopathy associated with deafness.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Bekesy G. Resting potentials inside the cochlear partition of guinea pig. Nature. 1952;169:241–242. - PubMed
-
- Hudspeth AJ. How the ear’s works work. Nature. 1989;341:397–404. - PubMed
-
- Davis H. Some principles of sensory receptor action. Physiol Rev. 1961;41:391–416. - PubMed
-
- Wangemann P, Schacht J. Homeostasic mechanisms in the cochlea. In: Dallos P, Popper AN, Fay R, editors. Springer Handbook of Auditory Research: The Cochlea. New York: Springer; 1996.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
