

Cellular retinoic acid-binding proteins are essential for hindbrain patterning and signal robustness in zebrafish
- PMID: 22619388
- PMCID: PMC3357909
- DOI: 10.1242/dev.077065
Cellular retinoic acid-binding proteins are essential for hindbrain patterning and signal robustness in zebrafish
Abstract
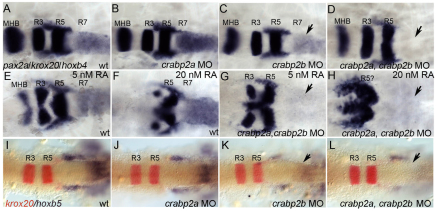
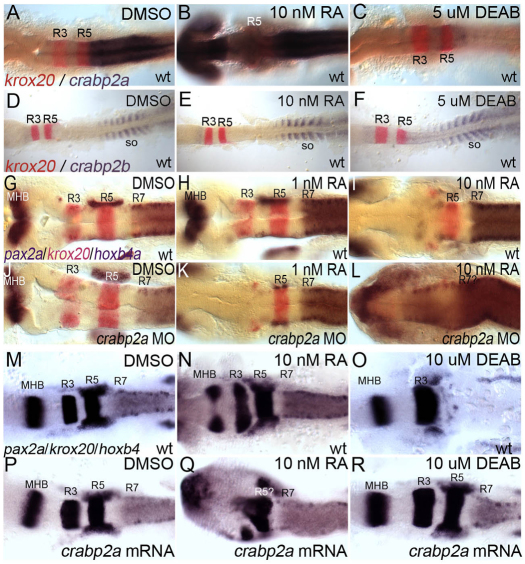
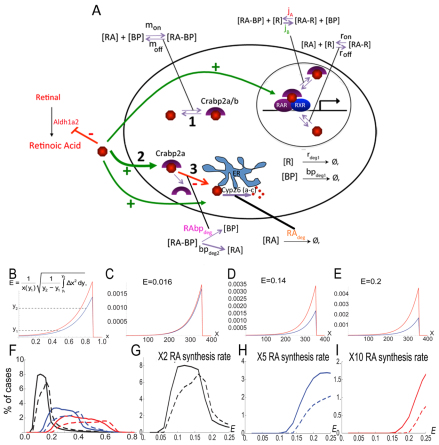
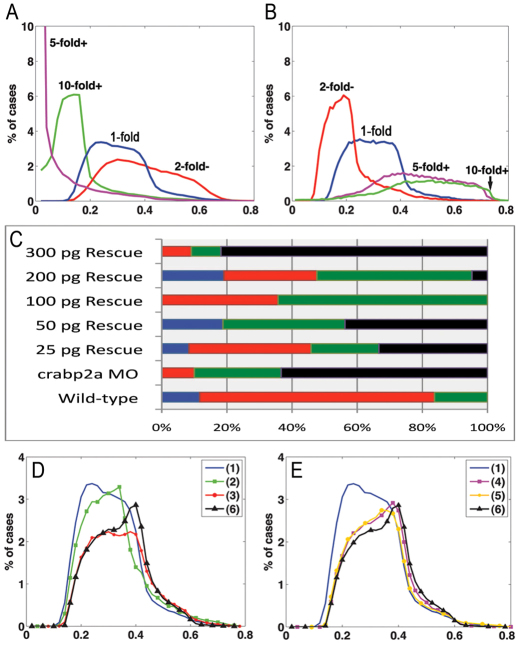
The vitamin A derivative retinoic acid (RA) is a morphogen that patterns the anterior-posterior axis of the vertebrate hindbrain. Cellular retinoic acid-binding proteins (Crabps) transport RA within cells to both its nuclear receptors (RARs) and degrading enzymes (Cyp26s). However, mice lacking Crabps are viable, suggesting that Crabp functions are redundant with those of other fatty acid-binding proteins. Here we show that Crabps in zebrafish are essential for posterior patterning of the hindbrain and that they provide a key feedback mechanism that makes signaling robust as they are able to compensate for changes in RA production. Of the four zebrafish Crabps, Crabp2a is uniquely RA inducible and depletion or overexpression of Crabp2a makes embryos hypersensitive to exogenous RA. Computational models confirm that Crabp2a improves robustness within a narrow concentration range that optimizes a 'robustness index', integrating spatial information along the RA morphogen gradient. Exploration of signaling parameters in our models suggests that the ability of Crabp2a to transport RA to Cyp26 enzymes for degradation is a major factor in promoting robustness. These results demonstrate a previously unrecognized requirement for Crabps in RA signaling and hindbrain development, as well as a novel mechanism for stabilizing morphogen gradients despite genetic or environmental fluctuations in morphogen availability.
Figures
References
-
- Allenby G., Janocha R., Kazmer S., Speck J., Grippo J. F., Levin A. A. (1994). Binding of 9-cis-retinoic acid and all-trans-retinoic acid to retinoic acid receptors alpha, beta, and gamma. Retinoic acid receptor gamma binds all-trans-retinoic acid preferentially over 9-cis-retinoic acid. J. Biol. Chem. 269, 16689–16695 - PubMed
-
- Aström A., Tavakkol A., Pettersson U., Cromie M., Elder J. T., Voorhees J. J. (1991). Molecular cloning of two human cellular retinoic acid-binding proteins (CRABP). Retinoic acid-induced expression of CRABP-II but not CRABP-I in adult human skin in vivo and in skin fibroblasts in vitro. J. Biol. Chem. 266, 17662–17666 - PubMed
-
- Aström A., Pettersson U., Chambon P., Voorhees J. J. (1994). Retinoic acid induction of human cellular retinoic acid-binding protein-II gene transcription is mediated by retinoic acid receptor-retinoid X receptor heterodimers bound to one far upstream retinoic acid-responsive element with 5-base pair spacing. J. Biol. Chem. 269, 22334–22339 - PubMed
-
- Begemann G., Schilling T. F., Rauch G. J., Geisler R., Ingham P. W. (2001). The zebrafish neckless mutation reveals a requirement for raldh2 in mesodermal signals that pattern the hindbrain. Development 128, 3081–3094 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
