

Blood flow reprograms lymphatic vessels to blood vessels
- PMID: 22622036
- PMCID: PMC3366395
- DOI: 10.1172/JCI57513
Blood flow reprograms lymphatic vessels to blood vessels
Erratum in
- J Clin Invest. 2012 Jul 2;122(7):2702. Vincente, Andres [corrected to Vicente, Andres]
Abstract
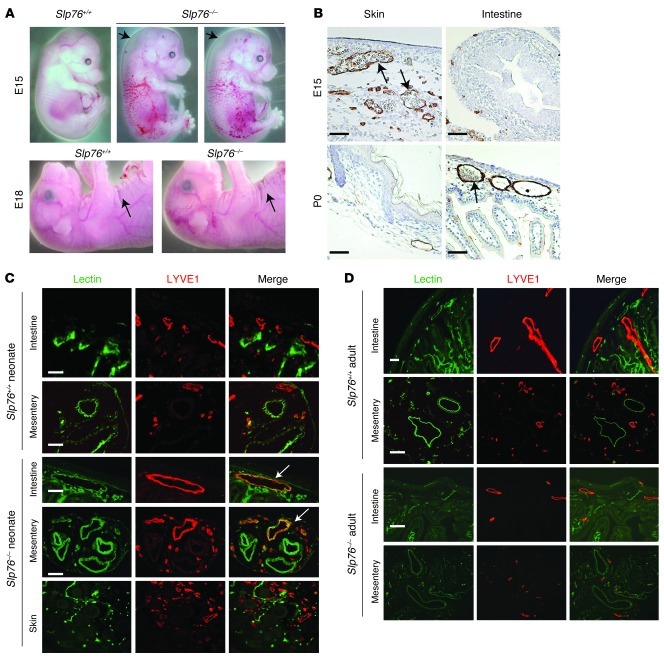
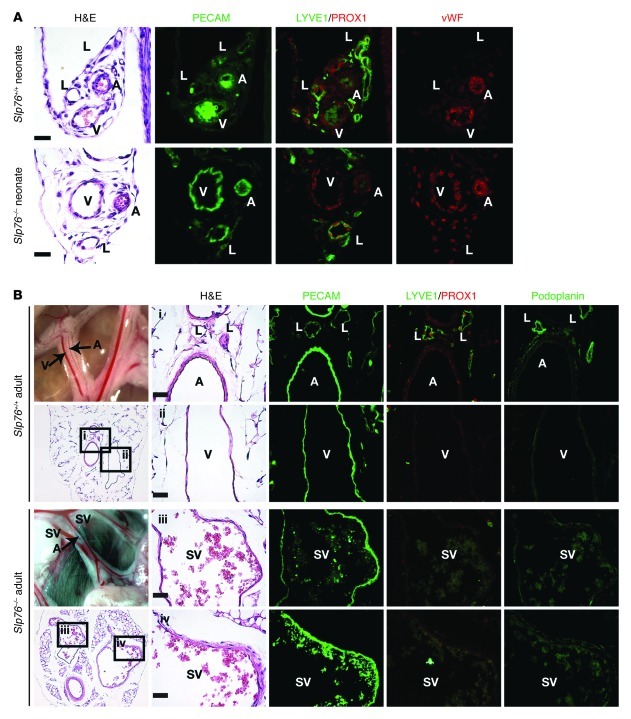
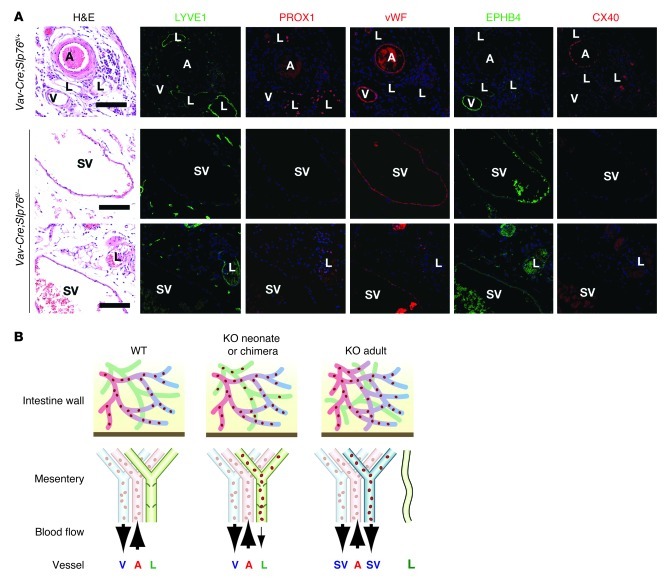
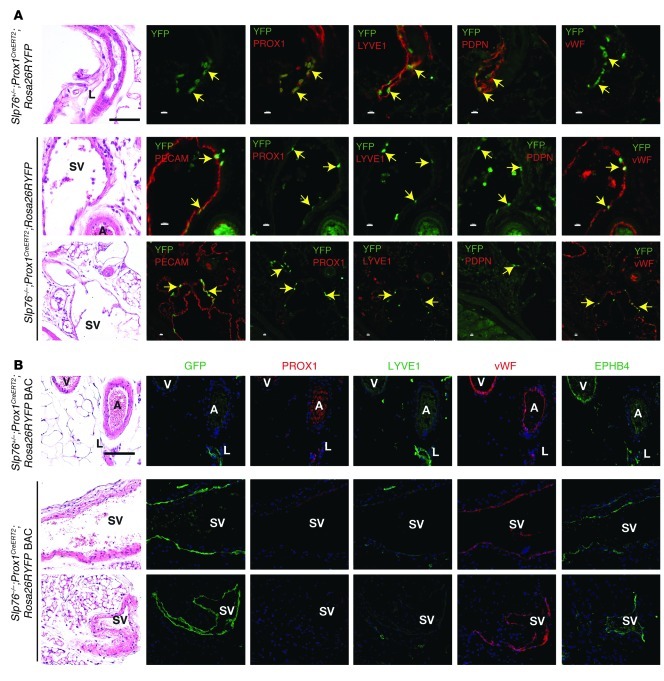
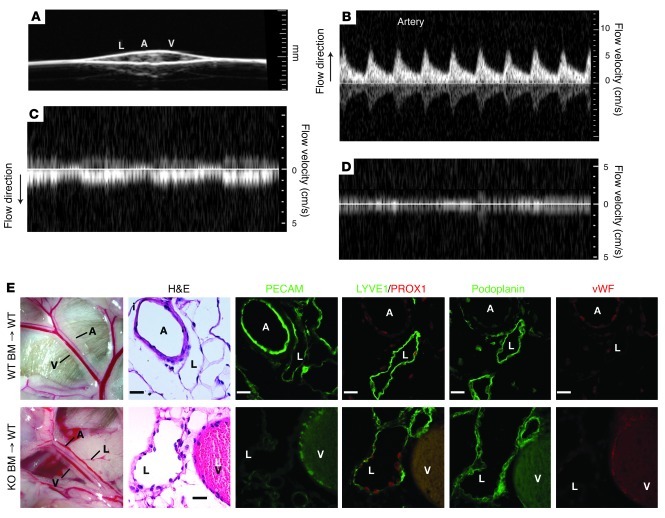
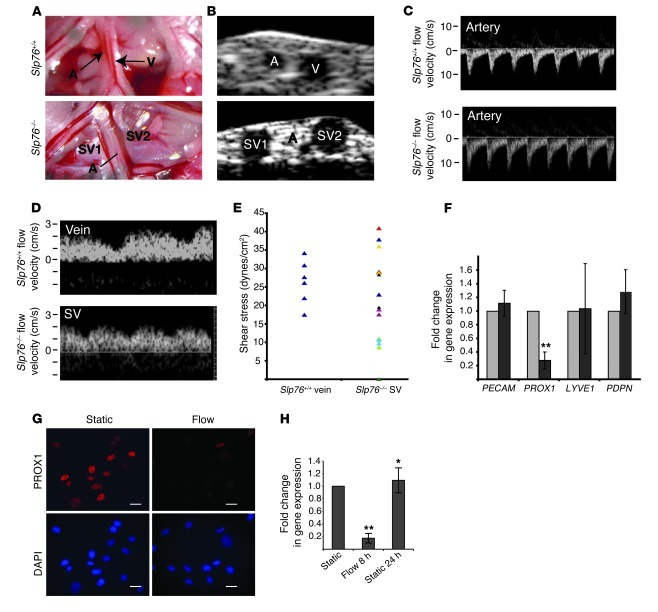
Human vascular malformations cause disease as a result of changes in blood flow and vascular hemodynamic forces. Although the genetic mutations that underlie the formation of many human vascular malformations are known, the extent to which abnormal blood flow can subsequently influence the vascular genetic program and natural history is not. Loss of the SH2 domain-containing leukocyte protein of 76 kDa (SLP76) resulted in a vascular malformation that directed blood flow through mesenteric lymphatic vessels after birth in mice. Mesenteric vessels in the position of the congenital lymphatic in mature Slp76-null mice lacked lymphatic identity and expressed a marker of blood vessel identity. Genetic lineage tracing demonstrated that this change in vessel identity was the result of lymphatic endothelial cell reprogramming rather than replacement by blood endothelial cells. Exposure of lymphatic vessels to blood in the absence of significant flow did not alter vessel identity in vivo, but lymphatic endothelial cells exposed to similar levels of shear stress ex vivo rapidly lost expression of PROX1, a lymphatic fate-specifying transcription factor. These findings reveal that blood flow can convert lymphatic vessels to blood vessels, demonstrating that hemodynamic forces may reprogram endothelial and vessel identity in cardiovascular diseases associated with abnormal flow.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
