

Sustained translational repression by eIF2α-P mediates prion neurodegeneration
- PMID: 22622579
- PMCID: PMC3378208
- DOI: 10.1038/nature11058
Sustained translational repression by eIF2α-P mediates prion neurodegeneration
Erratum in
- Nature.2014 Jul 17;511(7509):370
Abstract
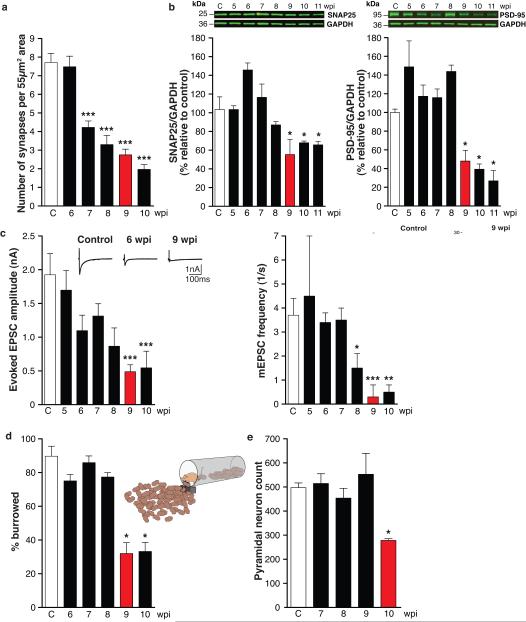
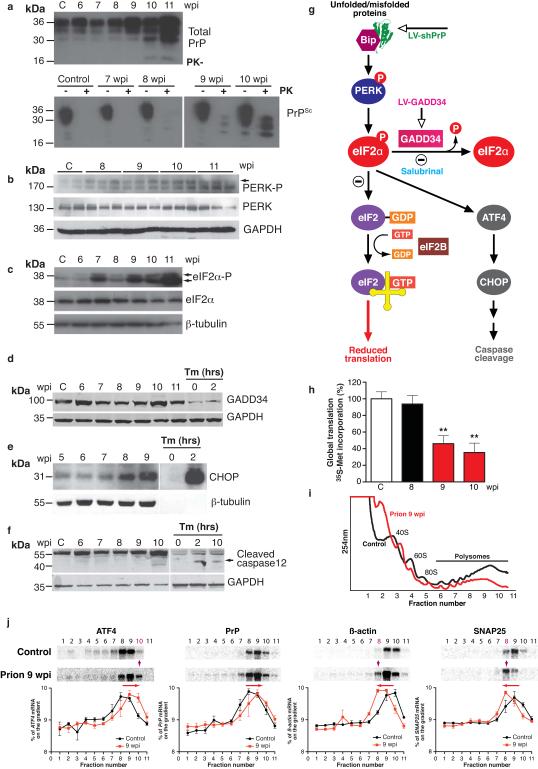
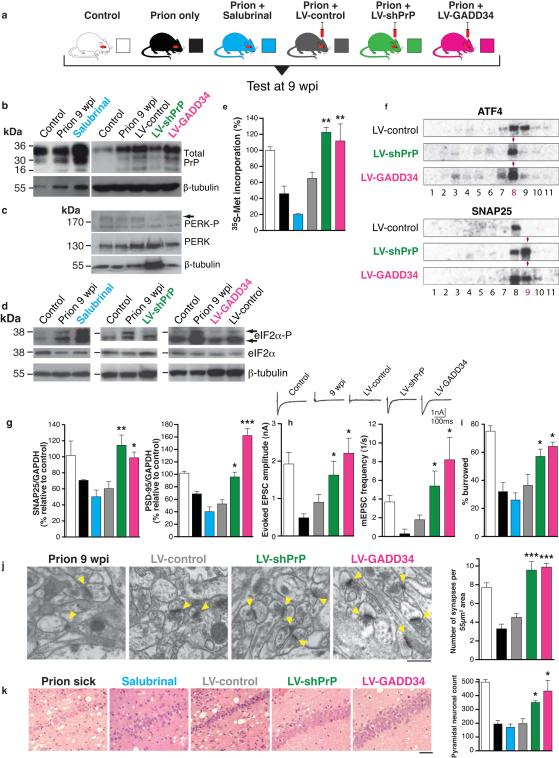
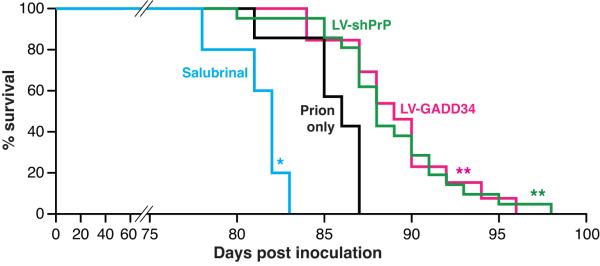
The mechanisms leading to neuronal death in neurodegenerative disease are poorly understood. Many of these disorders, including Alzheimer's, Parkinson's and prion diseases, are associated with the accumulation of misfolded disease-specific proteins. The unfolded protein response is a protective cellular mechanism triggered by rising levels of misfolded proteins. One arm of this pathway results in the transient shutdown of protein translation, through phosphorylation of the α-subunit of eukaryotic translation initiation factor, eIF2. Activation of the unfolded protein response and/or increased eIF2α-P levels are seen in patients with Alzheimer's, Parkinson's and prion diseases, but how this links to neurodegeneration is unknown. Here we show that accumulation of prion protein during prion replication causes persistent translational repression of global protein synthesis by eIF2α-P, associated with synaptic failure and neuronal loss in prion-diseased mice. Further, we show that promoting translational recovery in hippocampi of prion-infected mice is neuroprotective. Overexpression of GADD34, a specific eIF2α-P phosphatase, as well as reduction of levels of prion protein by lentivirally mediated RNA interference, reduced eIF2α-P levels. As a result, both approaches restored vital translation rates during prion disease, rescuing synaptic deficits and neuronal loss, thereby significantly increasing survival. In contrast, salubrinal, an inhibitor of eIF2α-P dephosphorylation, increased eIF2α-P levels, exacerbating neurotoxicity and significantly reducing survival in prion-diseased mice. Given the prevalence of protein misfolding and activation of the unfolded protein response in several neurodegenerative diseases, our results suggest that manipulation of common pathways such as translational control, rather than disease-specific approaches, may lead to new therapies preventing synaptic failure and neuronal loss across the spectrum of these disorders.
Figures
Comment in
-
Lost in translation: misfolded proteins may cause neurodegeneration by inhibiting normal protein production.Mov Disord. 2012 Sep 1;27(10):1218. doi: 10.1002/mds.25130. Mov Disord. 2012. PMID: 23136660 No abstract available.
References
-
- Hoozemans JJ, et al. Activation of the unfolded protein response in Parkinson’s disease. Biochem Biophys Res Commun. 2007;354:707–711. - PubMed
-
- Hoozemans JJ, et al. The unfolded protein response is activated in Alzheimer′s disease. Acta Neuropathol. 2005;110:165–172. - PubMed
-
- Unterberger U, et al. Endoplasmic reticulum stress features are prominent in Alzheimer disease but not in prion diseases in vivo. J Neuropathol Exp Neurol. 2006;65:348–357. - PubMed
-
- Boyce M, et al. A selective inhibitor of eIF2alpha dephosphorylation protects cells from ER stress. Science. 2005;307:935–939. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MC_U132681855/MRC_/Medical Research Council/United Kingdom
- U.1051.02.011.00001.01 (85860)/MRC_/Medical Research Council/United Kingdom
- MC_U132692719/MRC_/Medical Research Council/United Kingdom
- MC_UP_A600_1024/MRC_/Medical Research Council/United Kingdom
- MC_U123160654/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
