

Muenke syndrome mutation, FgfR3P²⁴⁴R, causes TMJ defects
- PMID: 22622662
- PMCID: PMC3383850
- DOI: 10.1177/0022034512449170
Muenke syndrome mutation, FgfR3P²⁴⁴R, causes TMJ defects
Abstract
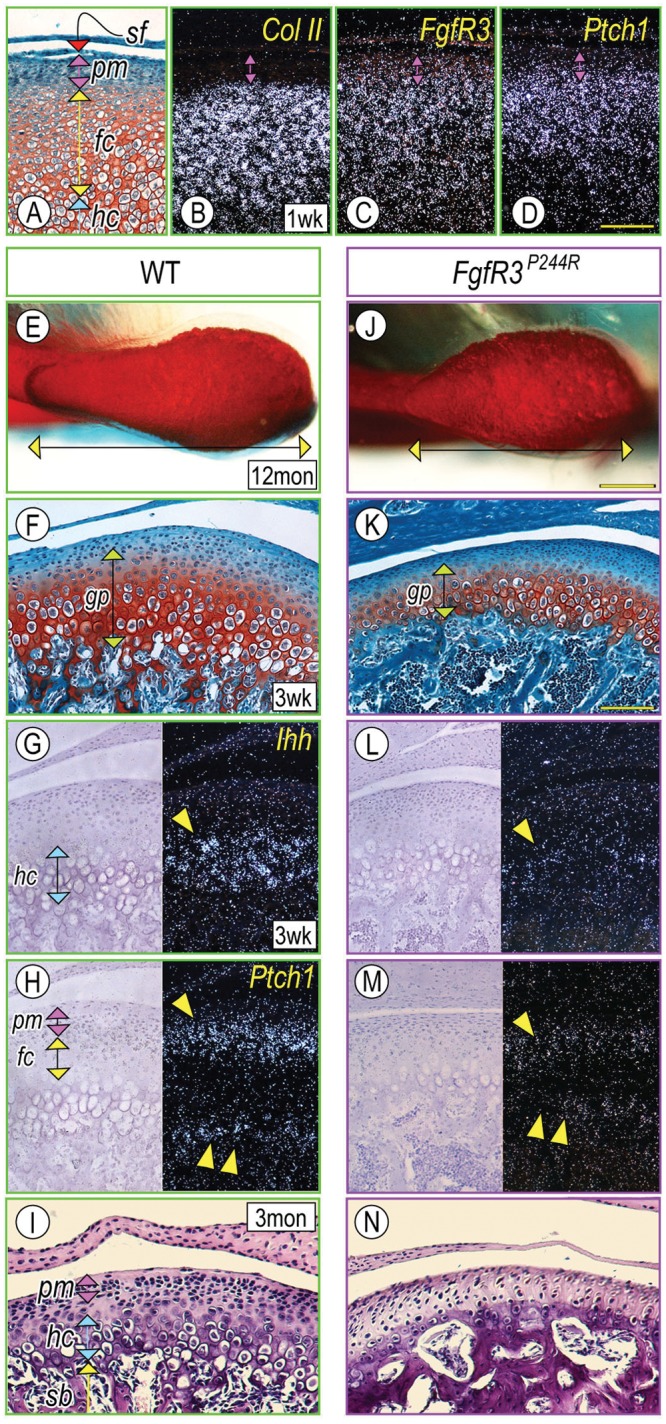
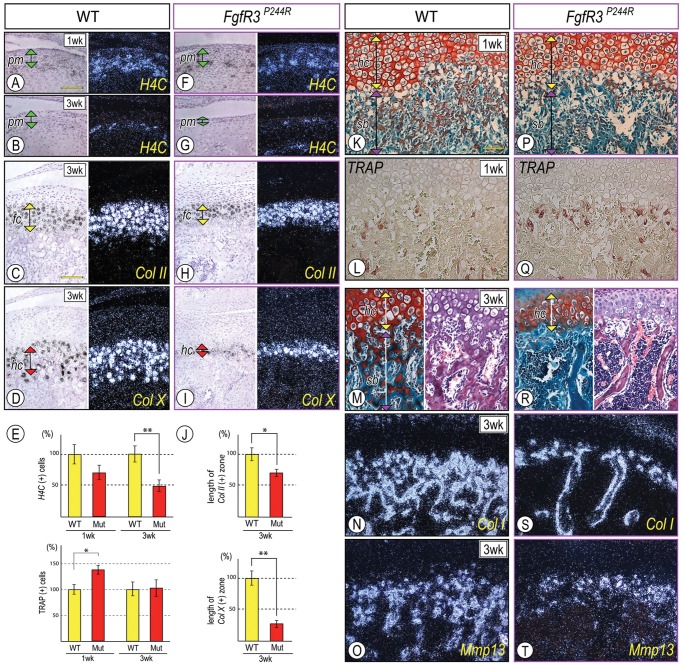
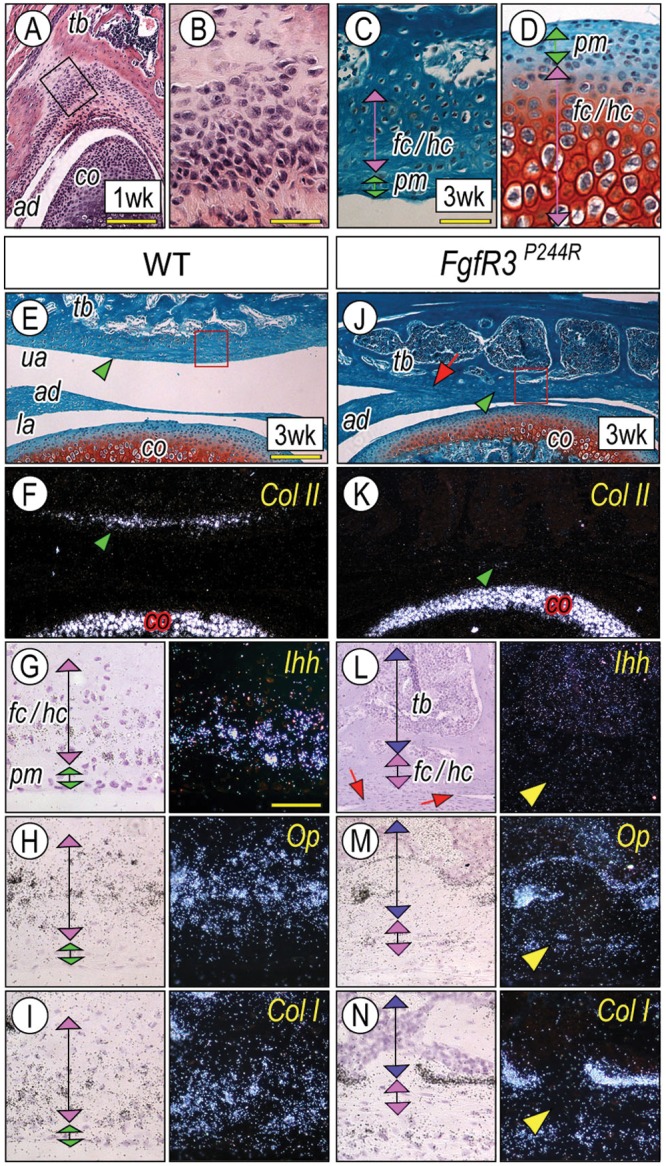
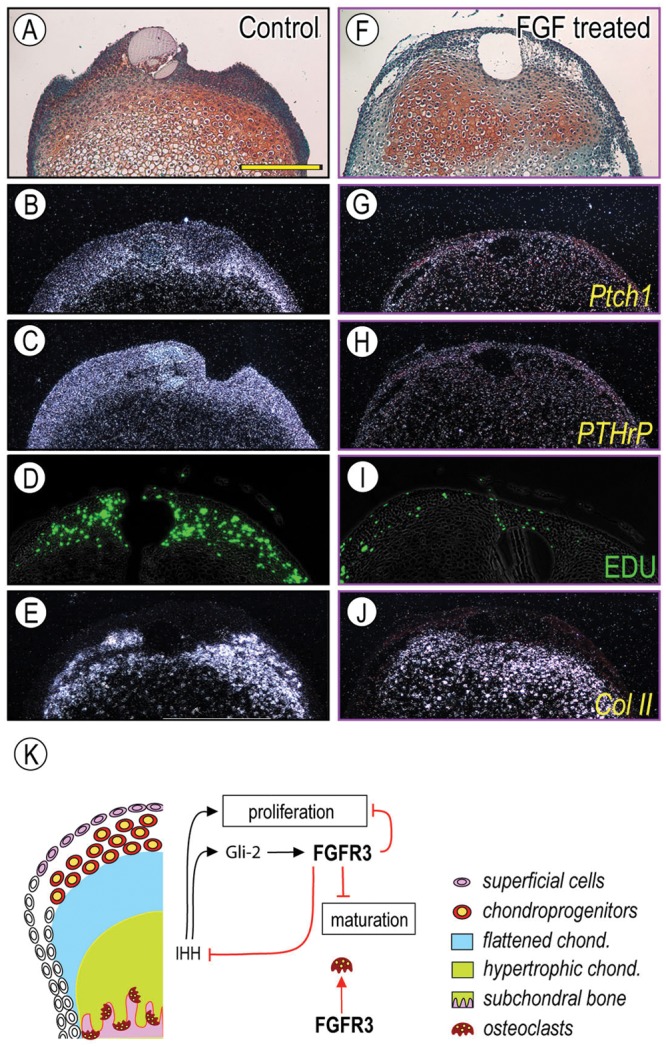
Muenke syndrome is characterized by various craniofacial deformities and is caused by an autosomal-dominant activating mutation in fibroblast growth factor receptor 3 (FGFR3(P250R) ). Here, using mice carrying a corresponding mutation (FgfR3(P244R) ), we determined whether the mutation affects temporomandibular joint (TMJ) development and growth. In situ hybridization showed that FgfR3 was expressed in condylar chondroprogenitors and maturing chondrocytes that also expressed the Indian hedgehog (Ihh) receptor and transcriptional target Patched 1(Ptch1). In FgfR3(P244R) mutants, the condyles displayed reduced levels of Ihh expression, H4C-positive proliferating chondroprogenitors, and collagen type II- and type X-expressing chondrocytes. Primary bone spongiosa formation was also disturbed and was accompanied by increased osteoclastic activity and reduced trabecular bone formation. Treatment of wild-type condylar explants with recombinant FGF2/FGF9 decreased Ptch1 and PTHrP expression in superficial/polymorphic layers and proliferation in chondroprogenitors. We also observed early degenerative changes of condylar articular cartilage, abnormal development of the articular eminence/glenoid fossa in the TMJ, and fusion of the articular disc. Analysis of our data indicates that the activating FgfR3(P244R) mutation disturbs TMJ developmental processes, likely by reducing hedgehog signaling and endochondral ossification. We suggest that a balance between FGF and hedgehog signaling pathways is critical for the integrity of TMJ development and for the maintenance of cellular organization.
Conflict of interest statement
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
Figures
References
-
- Colvin JS, Bohne BA, Harding GW, McEwen DG, Ornitz DM. (1996). Skeletal overgrowth and deafness in mice lacking fibroblast growth factor receptor 3. Nat Genet 12:390-397 - PubMed
-
- Deng C, Wynshaw-Boris A, Zhou F, Kuo A, Leder P. (1996). Fibroblast growth factor receptor 3 is a negative regulator of bone growth. Cell 84:911-921 - PubMed
-
- Hinton RJ, McNamara JA., Jr (1984). Temporal bone adaptations in response to protrusive function in juvenile and young adult rhesus monkeys (Macaca mulatta). Eur J Orthod 6:155-174 - PubMed
-
- Ibrahimi OA, Zhang F, Eliseenkova AV, Linhardt RJ, Mohammadi M. (2004). Proline to arginine mutations in FGF receptors 1 and 3 result in Pfeiffer and Muenke craniosynostosis syndromes through enhancement of FGF binding affinity. Hum Mol Genet 13:69-78 - PubMed
-
- Kantomaa T, Tuominen M, Pirttiniemi P. (1994). Effect of mechanical forces on chondrocyte maturation and differentiation in the mandibular condyle of the rat. J Dent Res 73:1150-1156 - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
