

In vivo visualization of delta opioid receptors upon physiological activation uncovers a distinct internalization profile
- PMID: 22623675
- PMCID: PMC3371401
- DOI: 10.1523/JNEUROSCI.0185-12.2012
In vivo visualization of delta opioid receptors upon physiological activation uncovers a distinct internalization profile
Abstract
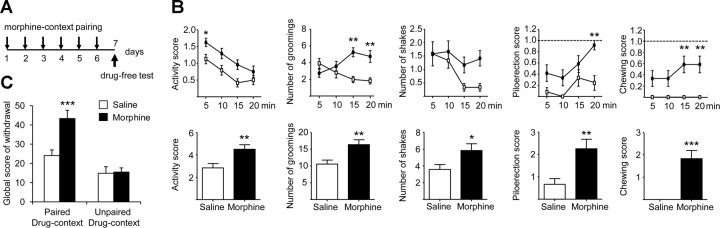
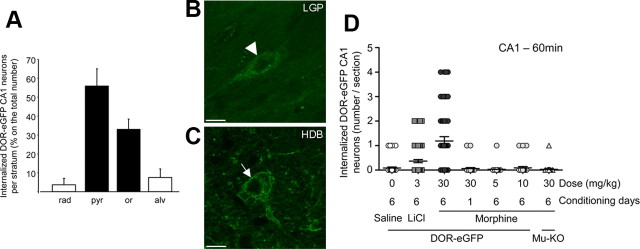
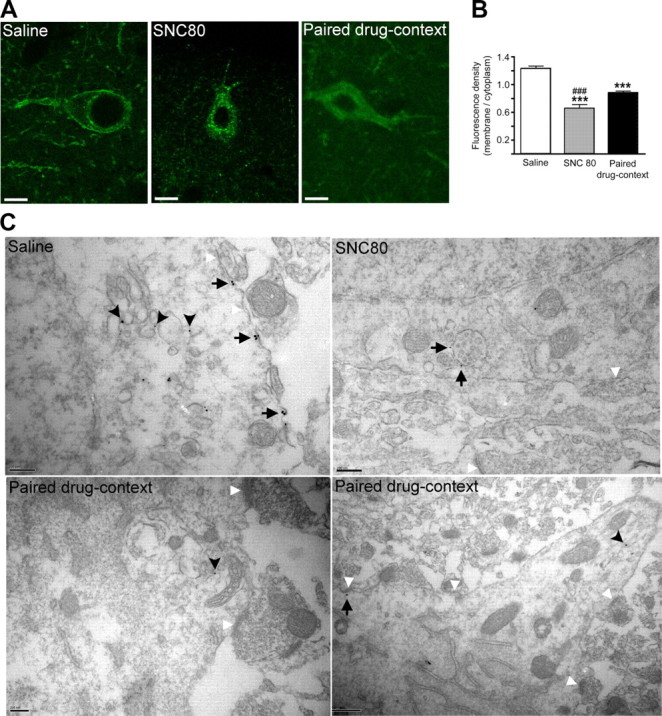
G-protein-coupled receptors (GPCRs) mediate numerous physiological functions and represent prime therapeutic targets. Receptor trafficking upon agonist stimulation is critical for GPCR function, but examining this process in vivo remains a true challenge. Using knock-in mice expressing functional fluorescent delta opioid receptors under the control of the endogenous promoter, we visualized in vivo internalization of this native GPCR upon physiological stimulation. We developed a paradigm in which animals were made dependent on morphine in a drug-paired context. When re-exposed to this context in a drug-free state, mice showed context-dependent withdrawal signs and activation of the hippocampus. Receptor internalization was transiently detected in a subset of CA1 neurons, uncovering regionally restricted opioid peptide release. Importantly, a pool of surface receptors always remained, which contrasts with the in vivo profile previously established for exogenous drug-induced internalization. Therefore, a distinct response is observed at the receptor level upon a physiological or pharmacological stimulation. Altogether, direct in vivo GPCR visualization enables mapping receptor stimulation promoted by a behavioral challenge and represents a powerful approach to study endogenous GPCR physiology.
Figures






Similar articles
-
Impact of chronic morphine on delta opioid receptor-expressing neurons in the mouse hippocampus.Neuroscience. 2016 Jan 28;313:46-56. doi: 10.1016/j.neuroscience.2015.10.022. Epub 2015 Oct 19. Neuroscience. 2016. PMID: 26480813 Free PMC article.
-
In vivo delta opioid receptor internalization controls behavioral effects of agonists.PLoS One. 2009;4(5):e5425. doi: 10.1371/journal.pone.0005425. Epub 2009 May 1. PLoS One. 2009. PMID: 19412545 Free PMC article.
-
In vivo techniques to investigate the internalization profile of opioid receptors.Methods Mol Biol. 2015;1230:87-104. doi: 10.1007/978-1-4939-1708-2_7. Methods Mol Biol. 2015. PMID: 25293318 Free PMC article.
-
Role of delivery and trafficking of delta-opioid peptide receptors in opioid analgesia and tolerance.Trends Pharmacol Sci. 2006 Jun;27(6):324-9. doi: 10.1016/j.tips.2006.04.005. Epub 2006 May 6. Trends Pharmacol Sci. 2006. PMID: 16678916 Review.
-
The delta opioid receptor tool box.Neuroscience. 2016 Dec 3;338:145-159. doi: 10.1016/j.neuroscience.2016.06.028. Epub 2016 Jun 24. Neuroscience. 2016. PMID: 27349452 Free PMC article. Review.
Cited by
-
Sensing of amino acids by the gut-expressed taste receptor T1R1-T1R3 stimulates CCK secretion.Am J Physiol Gastrointest Liver Physiol. 2013 Feb 1;304(3):G271-82. doi: 10.1152/ajpgi.00074.2012. Epub 2012 Nov 29. Am J Physiol Gastrointest Liver Physiol. 2013. PMID: 23203156 Free PMC article.
-
The multiple facets of opioid receptor function: implications for addiction.Curr Opin Neurobiol. 2013 Aug;23(4):473-9. doi: 10.1016/j.conb.2013.02.005. Epub 2013 Feb 28. Curr Opin Neurobiol. 2013. PMID: 23453713 Free PMC article. Review.
-
Mapping Astrocytic and Neuronal μ-opioid Receptor Expression in Various Brain Regions Using MOR-mCherry Reporter Mouse.Exp Neurobiol. 2023 Dec 31;32(6):395-409. doi: 10.5607/en23039. Exp Neurobiol. 2023. PMID: 38196135 Free PMC article.
-
Strategy for Identification of Phosphorylation Levels of Low Abundance Proteins in Vivo for Which Antibodies Are not Available.J Cardiovasc Dev Dis. 2017 Oct 8;4(4):17. doi: 10.3390/jcdd4040017. J Cardiovasc Dev Dis. 2017. PMID: 29367546 Free PMC article.
-
Anti-nociception mediated by a κ opioid receptor agonist is blocked by a δ receptor agonist.Br J Pharmacol. 2015 Jan;172(2):691-703. doi: 10.1111/bph.12810. Epub 2014 Sep 5. Br J Pharmacol. 2015. PMID: 24923251 Free PMC article.
References
-
- Berrendero F, Castañé A, Ledent C, Parmentier M, Maldonado R, Valverde O. Increase of morphine withdrawal in mice lacking A2a receptors and no changes in CB1/A2a double knockout mice. Eur J Neurosci. 2003;17:315–324. - PubMed
-
- Ciccocioppo R, Martin-Fardon R, Weiss F. Effect of selective blockade of mu(1) or delta opioid receptors on reinstatement of alcohol-seeking behavior by drug-associated stimuli in rats. Neuropsychopharmacology. 2002;27:391–399. - PubMed
-
- David V, Matifas A, Gavello-Baudy S, Decorte L, Kieffer BL, Cazala P. Brain regional Fos expression elicited by the activation of mu- but not delta-opioid receptors of the ventral tegmental area: evidence for an implication of the ventral thalamus in opiate reward. Neuropsychopharmacology. 2008;33:1746–1759. - PubMed
-
- Decossas M, Bloch B, Bernard V. Trafficking of the muscarinic m2 autoreceptor in cholinergic basalocortical neurons in vivo: differential regulation of plasma membrane receptor availability and intraneuronal localization in acetylcholinesterase-deficient and -inhibited mice. J Comp Neurol. 2003;462:302–314. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous