

Running speed alters the frequency of hippocampal gamma oscillations
- PMID: 22623683
- PMCID: PMC3366345
- DOI: 10.1523/JNEUROSCI.5110-11.2012
Running speed alters the frequency of hippocampal gamma oscillations
Abstract
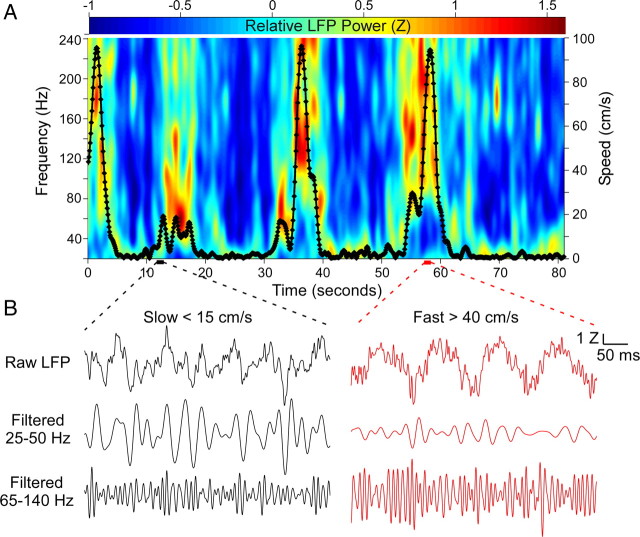
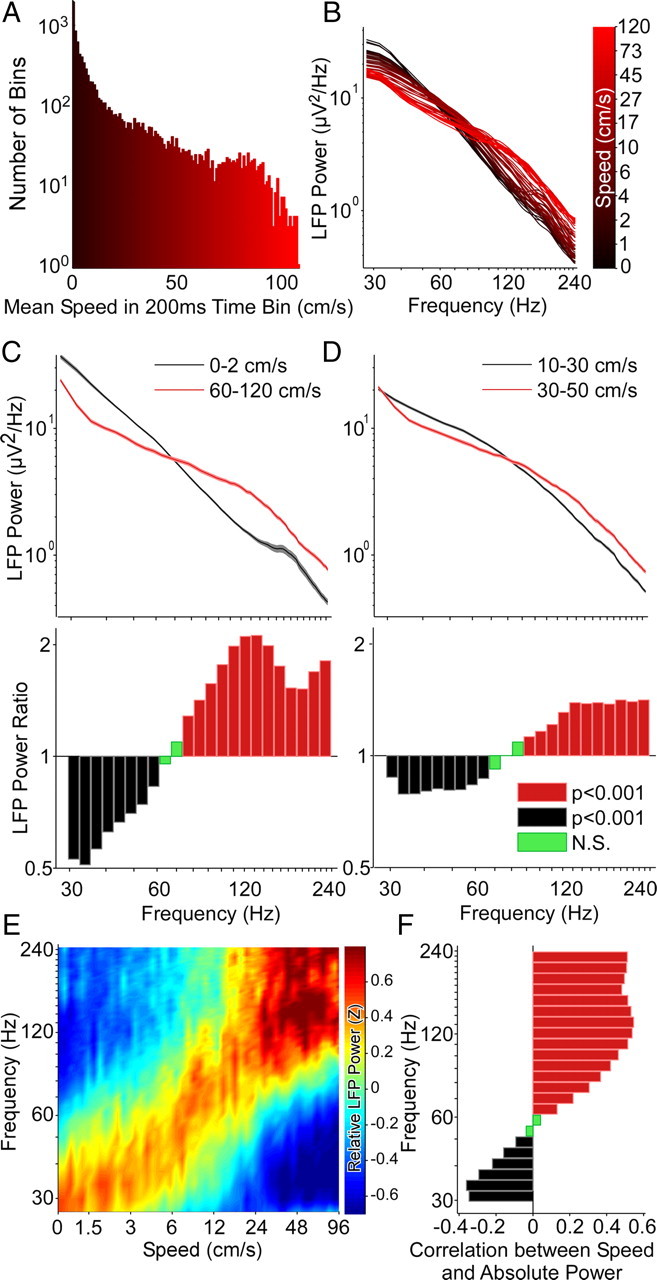
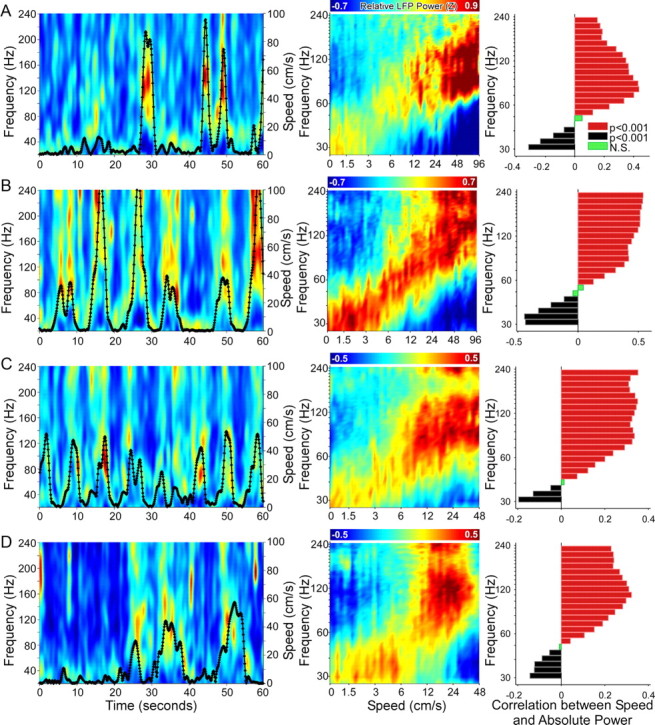
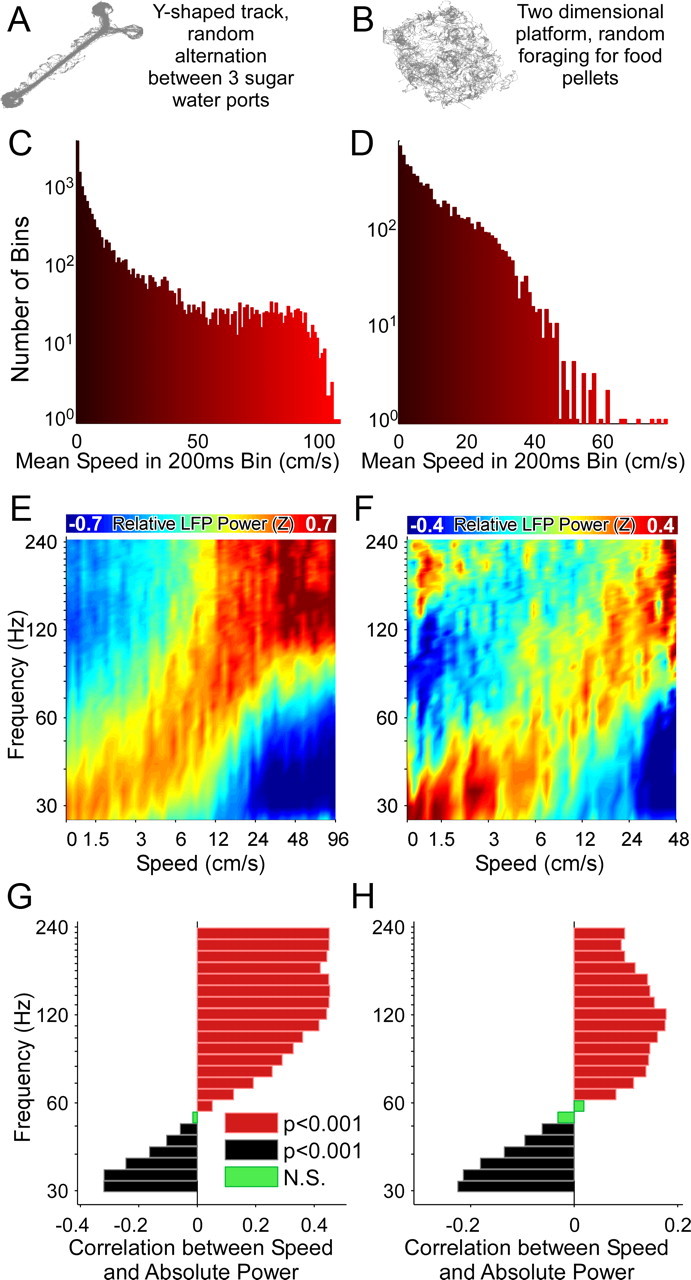
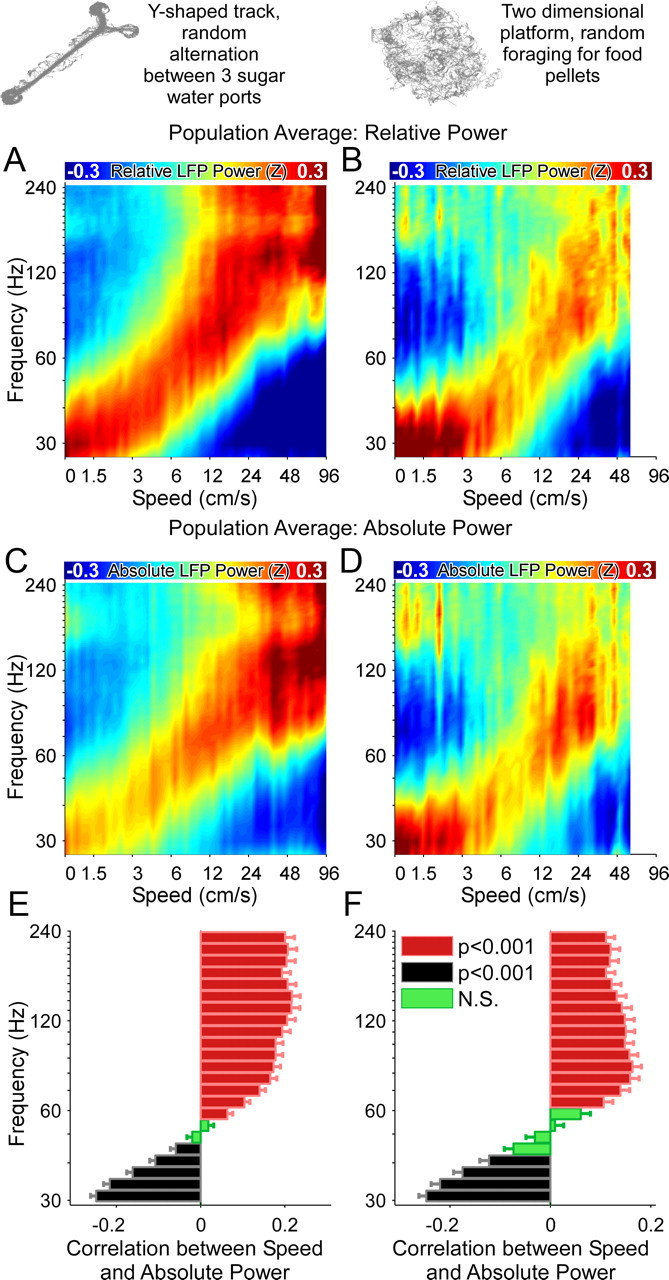
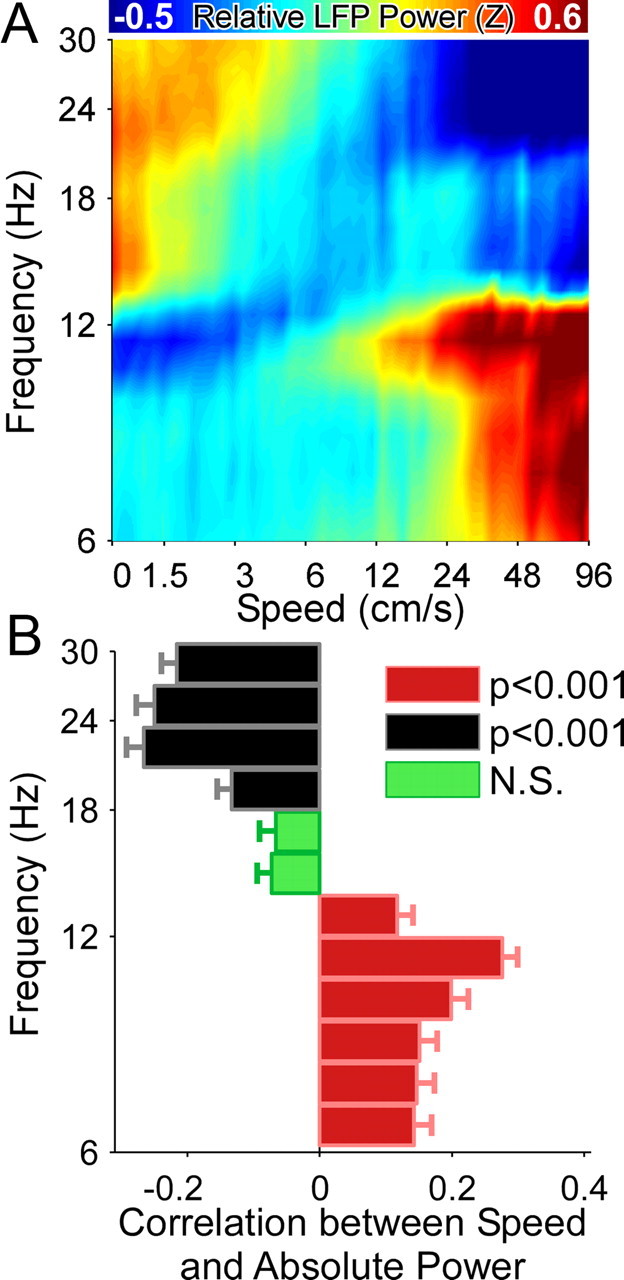
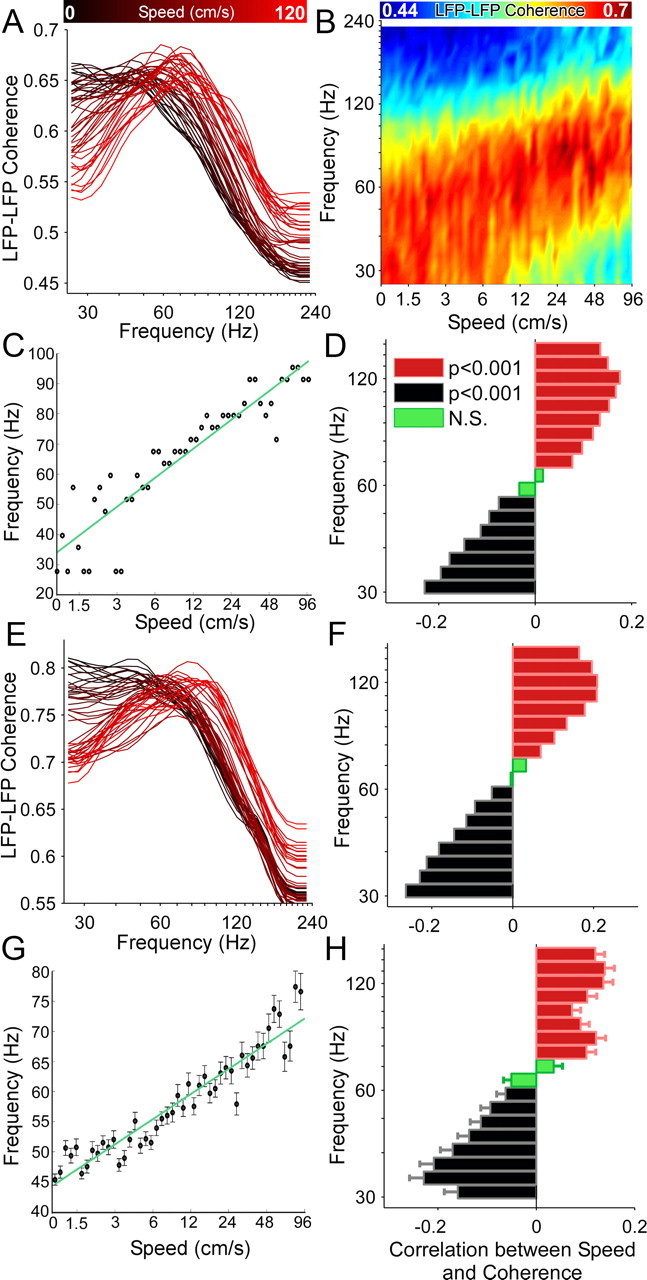
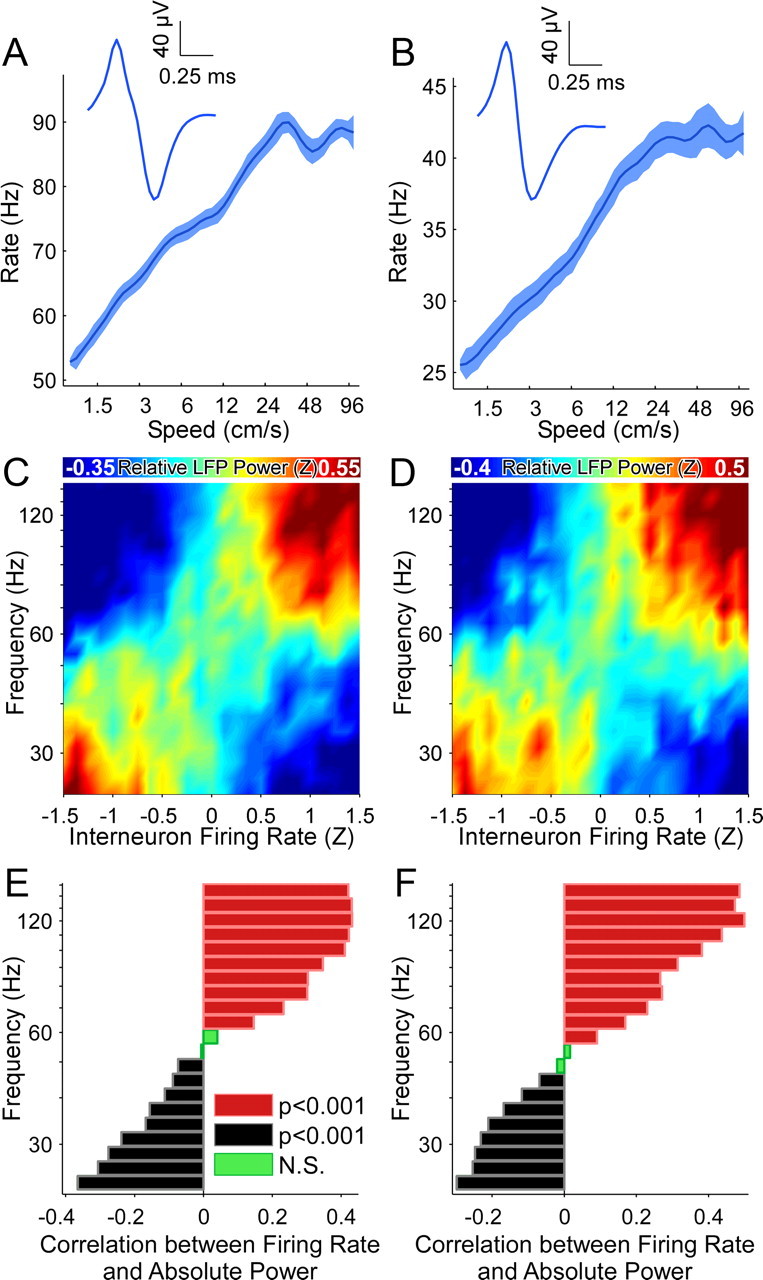
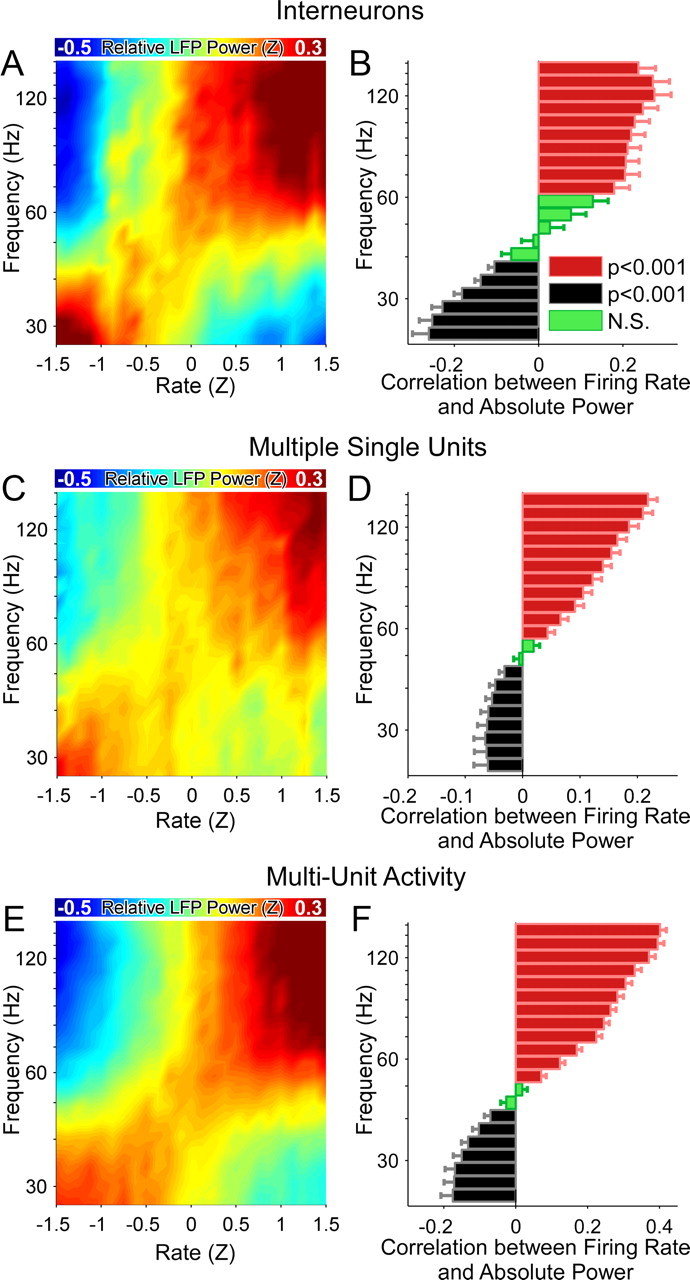
Successful spatial navigation is thought to employ a combination of at least two strategies: the following of landmark cues and path integration. Path integration requires that the brain use the speed and direction of movement in a meaningful way to continuously compute the position of the animal. Indeed, the running speed of rats modulates both the firing rate of neurons and the spectral properties of low frequency, theta oscillations seen in the local field potential (LFP) of the hippocampus, a region important for spatial memory formation. Higher frequency, gamma-band LFP oscillations are usually associated with decision-making, increased attention, and improved reaction times. Here, we show that increased running speed is accompanied by large, systematic increases in the frequency of hippocampal CA1 network oscillations spanning the entire gamma range (30-120 Hz) and beyond. These speed-dependent changes in frequency are seen on both linear tracks and two-dimensional platforms, and are thus independent of the behavioral task. Synchrony between anatomically distant CA1 regions also shifts to higher gamma frequencies as running speed increases. The changes in frequency are strongly correlated with changes in the firing rates of individual interneurons, consistent with models of gamma generation. Our results suggest that as a rat runs faster, there are faster gamma frequency transitions between sequential place cell-assemblies. This may help to preserve the spatial specificity of place cells and spatial memories at vastly different running speeds.
Figures
References
-
- Ahmed OJ. Brown University; 2010. Interpreting the rhythms of the hippocampus and neocortex. PhD thesis.
-
- Angelaki DE, Cullen KE. Vestibular system: the many facets of a multimodal sense. Annu Rev Neurosci. 2008;31:125–150. - PubMed
-
- Barlow JS. Inertial navigation as a basis for animal navigation. J Theor Biol. 1964;6:76–117. - PubMed
-
- Best PJ, White AM, Minai A. Spatial processing in the brain: the activity of hippocampal place cells. Annu Rev Neurosci. 2001;24:459–486. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous