

Neurogliaform and Ivy Cells: A Major Family of nNOS Expressing GABAergic Neurons
- PMID: 22623913
- PMCID: PMC3353154
- DOI: 10.3389/fncir.2012.00023
Neurogliaform and Ivy Cells: A Major Family of nNOS Expressing GABAergic Neurons
Abstract
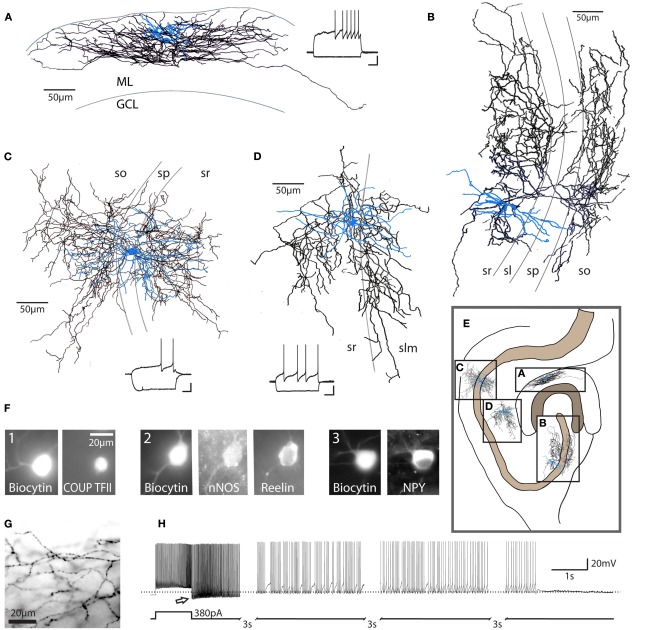
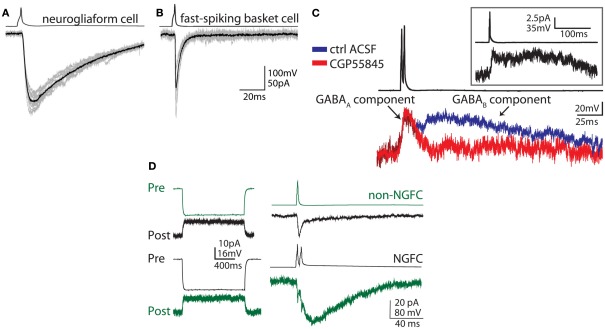
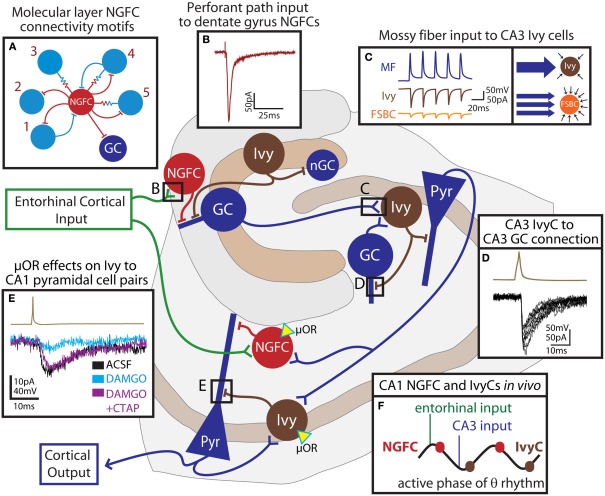
Neurogliaform and Ivy cells are members of an abundant family of neuronal nitric oxide synthase (nNOS) expressing GABAergic interneurons found in diverse brain regions. These cells have a defining dense local axonal plexus, and display unique synaptic properties including a biphasic postsynaptic response with both a slow GABA(A) component and a GABA(B) component following even a single action potential. The type of transmission displayed by these cells has been termed "volume transmission," distinct from both tonic and classical synaptic transmission. Electrical connections are also notable in that, unlike other GABAergic cell types, neurogliaform family cells will form gap junctions not only with other neurogliaform cells, but also with non-neurogliaform family GABAergic cells. In this review, we focus on neurogliaform and Ivy cells throughout the hippocampal formation, where recent studies highlight their role in feedforward inhibition, uncover their ability to display a phenomenon called persistent firing, and reveal their modulation by opioids. The unique properties of this family of cells, their abundance, rich connectivity, and modulation by clinically relevant drugs make them an attractive target for future studies in vivo during different behavioral and pharmacological conditions.
Keywords: GABAA,slow; GABAB; feedforward inhibition; opioid; persistent firing.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous