

Engineering rotor ring stoichiometries in the ATP synthase
- PMID: 22628564
- PMCID: PMC3382517
- DOI: 10.1073/pnas.1120027109
Engineering rotor ring stoichiometries in the ATP synthase
Abstract
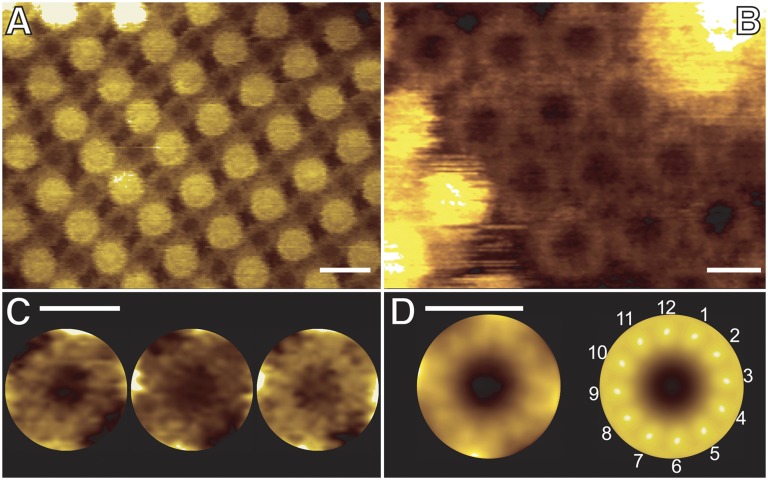
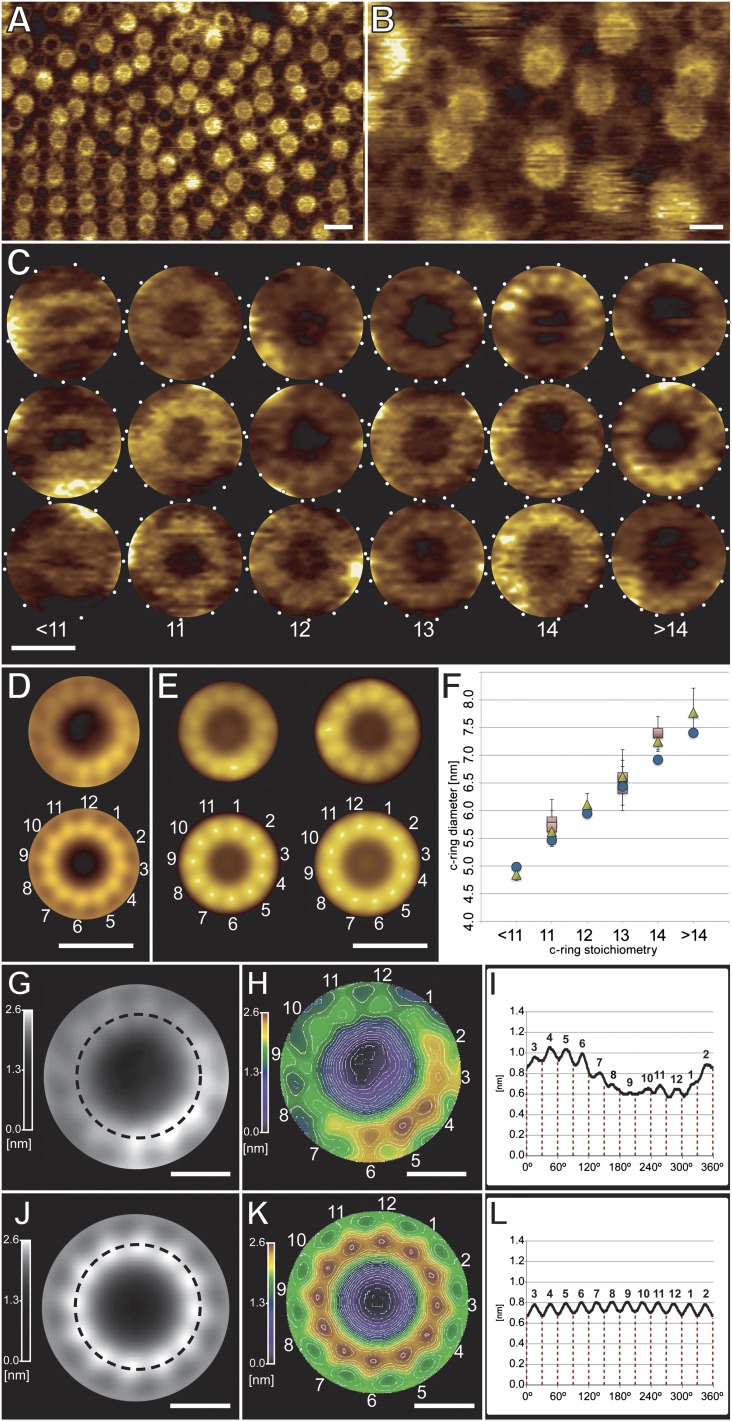
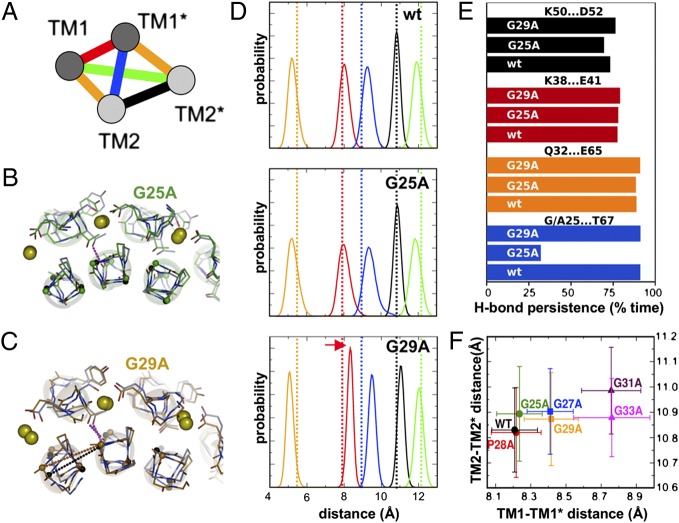
ATP synthase membrane rotors consist of a ring of c-subunits whose stoichiometry is constant for a given species but variable across different ones. We investigated the importance of c/c-subunit contacts by site-directed mutagenesis of a conserved stretch of glycines (GxGxGxGxG) in a bacterial c(11) ring. Structural and biochemical studies show a direct, specific influence on the c-subunit stoichiometry, revealing c(<11), c(12), c(13), c(14), and c(>14) rings. Molecular dynamics simulations rationalize this effect in terms of the energetics and geometry of the c-subunit interfaces. Quantitative data from a spectroscopic interaction study demonstrate that the complex assembly is independent of the c-ring size. Real-time ATP synthesis experiments in proteoliposomes show the mutant enzyme, harboring the larger c(12) instead of c(11), is functional at lower ion motive force. The high degree of compliance in the architecture of the ATP synthase rotor offers a rationale for the natural diversity of c-ring stoichiometries, which likely reflect adaptations to specific bioenergetic demands. These results provide the basis for bioengineering ATP synthases with customized ion-to-ATP ratios, by sequence modifications.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
-
- Abrahams JP, Leslie AGW, Lutter R, Walker JE. Structure at 2.8 Å resolution of F1-ATPase from bovine heart mitochondria. Nature. 1994;370:621–628. - PubMed
-
- von Ballmoos C, Wiedenmann A, Dimroth P. Essentials for ATP synthesis by F1F0 ATP synthases. Annu Rev Biochem. 2009;78:649–672. - PubMed
-
- Junge W, Sielaff H, Engelbrecht S. Torque generation and elastic power transmission in the rotary FoF1-ATPase. Nature. 2009;459:364–370. - PubMed
-
- Noji H, Yasuda R, Yoshida M, Kinosita K., Jr Direct observation of the rotation of F1-ATPase. Nature. 1997;386:299–302. - PubMed
-
- Iino R, Rondelez Y, Yoshida M, Noji H. Chemomechanical coupling in single-molecule F-type ATP synthase. J Bioenerg Biomembr. 2005;37:451–454. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
