

Evolution of plant p-type ATPases
- PMID: 22629273
- PMCID: PMC3355532
- DOI: 10.3389/fpls.2012.00031
Evolution of plant p-type ATPases
Abstract
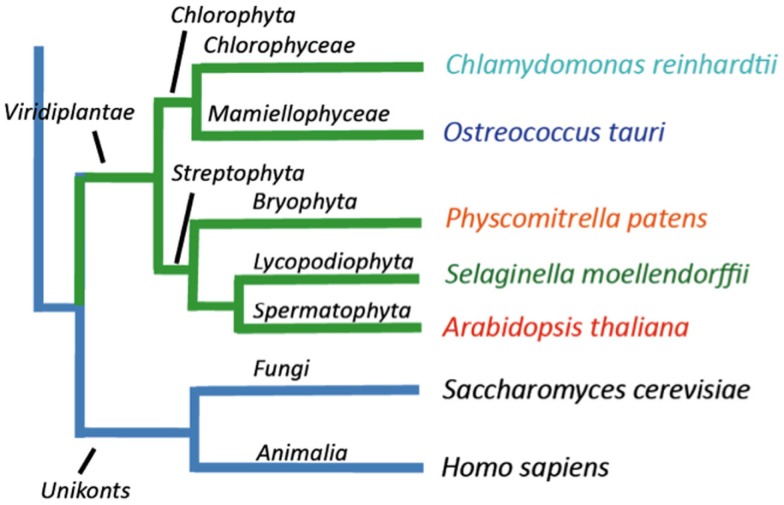
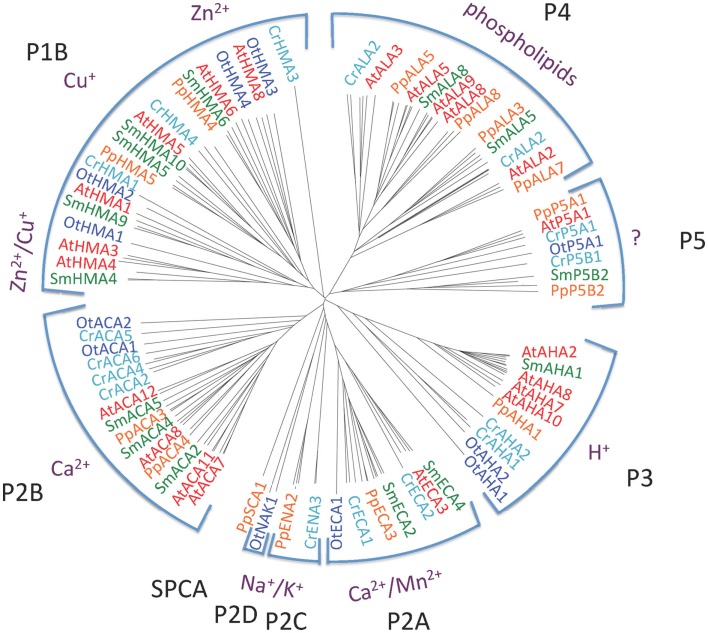
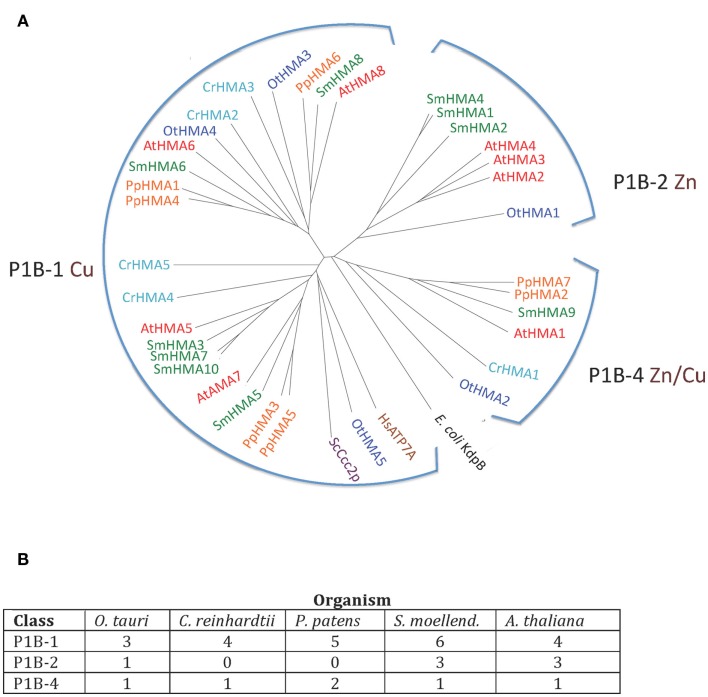
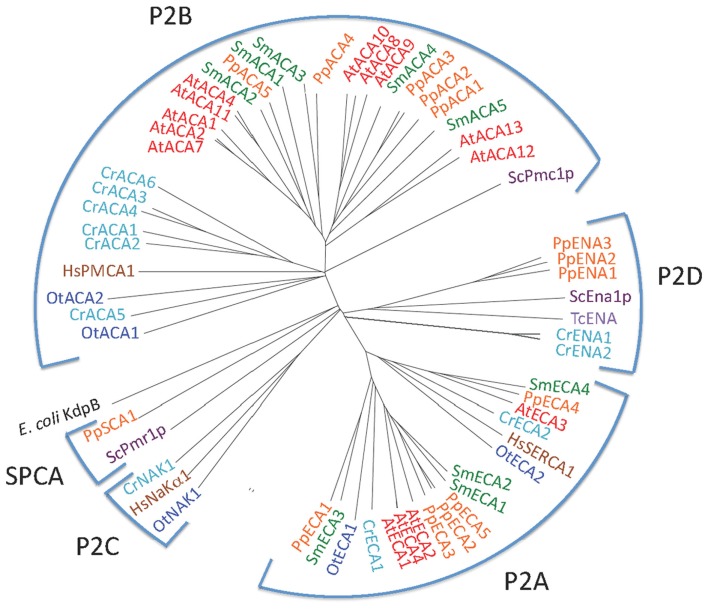
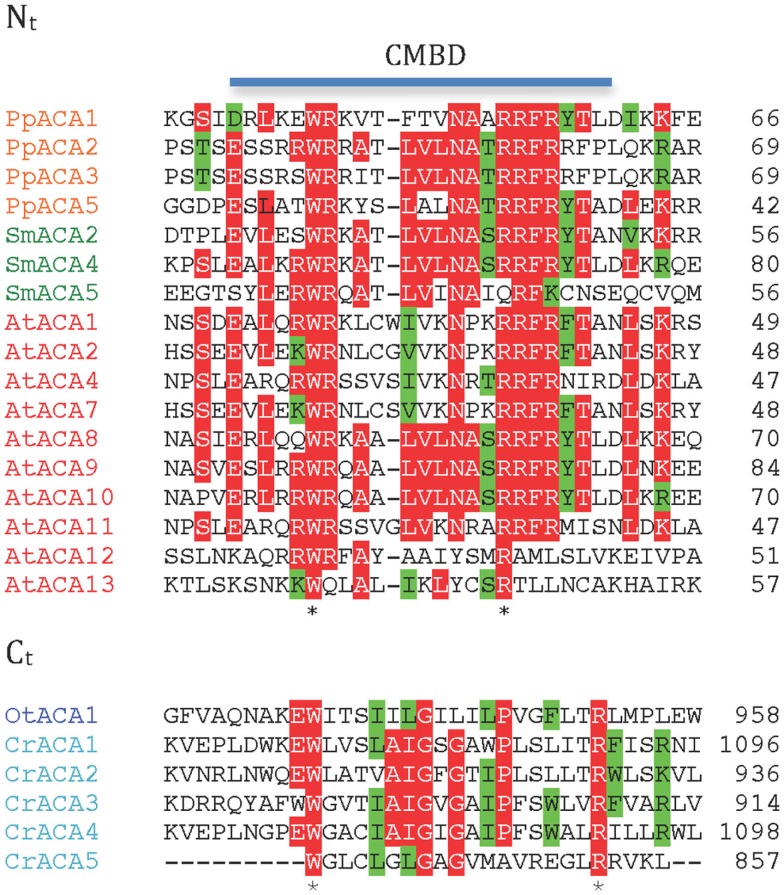
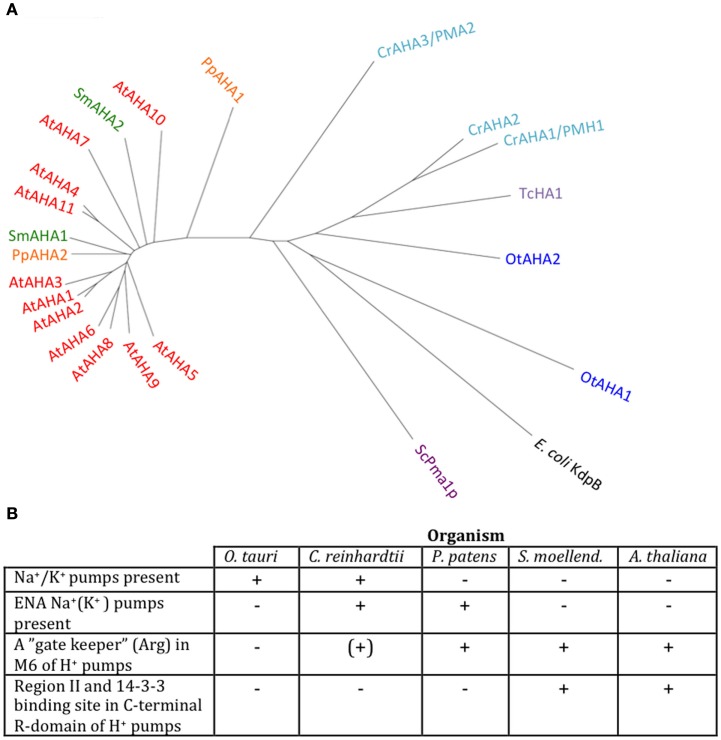
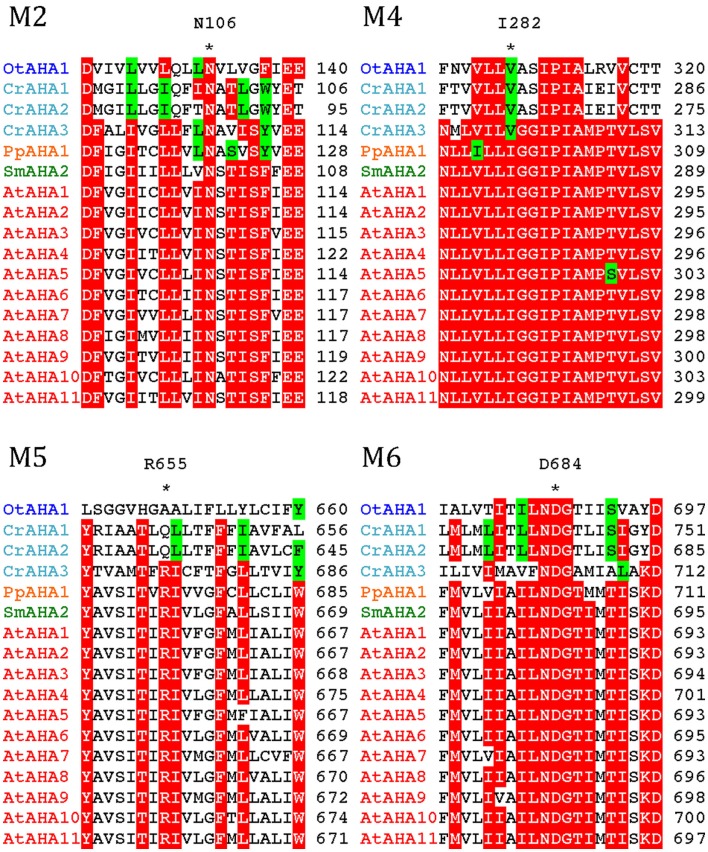
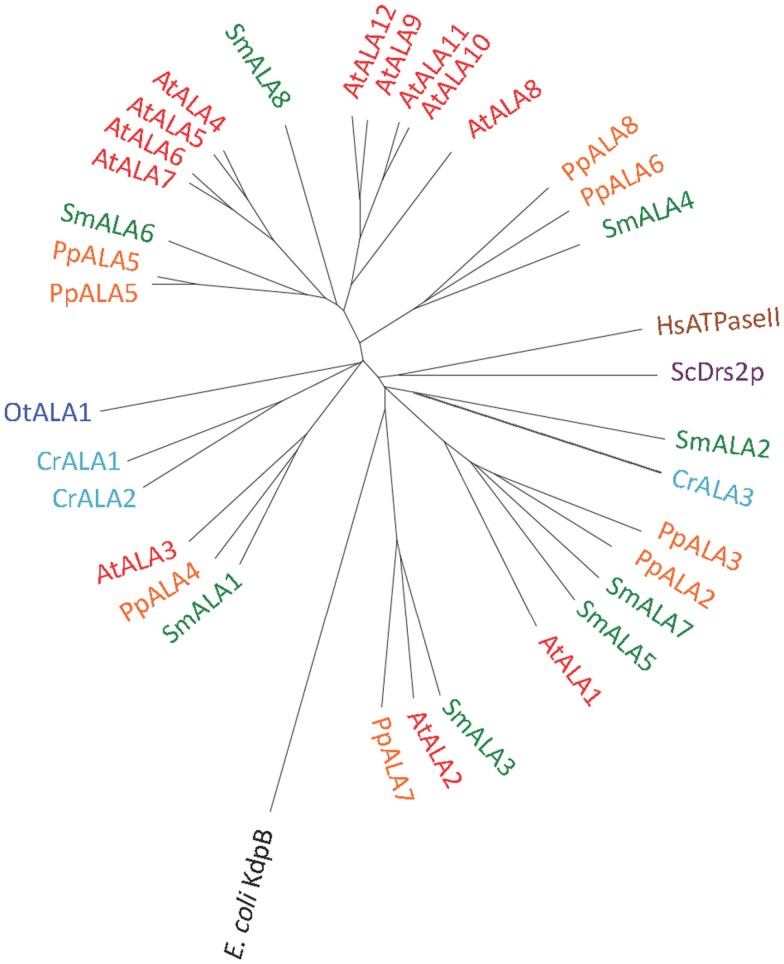
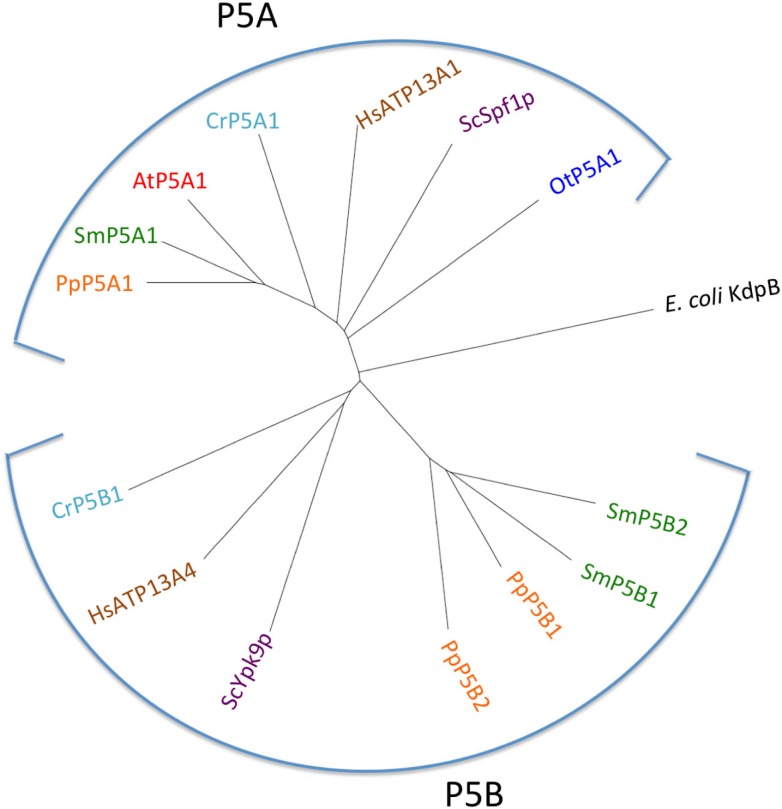
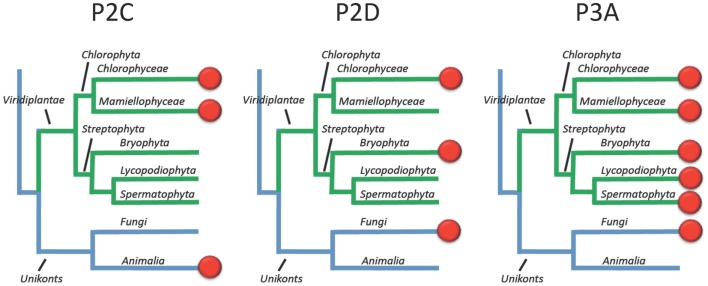
Five organisms having completely sequenced genomes and belonging to all major branches of green plants (Viridiplantae) were analyzed with respect to their content of P-type ATPases encoding genes. These were the chlorophytes Ostreococcus tauri and Chlamydomonas reinhardtii, and the streptophytes Physcomitrella patens (a non-vascular moss), Selaginella moellendorffii (a primitive vascular plant), and Arabidopsis thaliana (a model flowering plant). Each organism contained sequences for all five subfamilies of P-type ATPases. Whereas Na(+) and H(+) pumps seem to mutually exclude each other in flowering plants and animals, they co-exist in chlorophytes, which show representatives for two kinds of Na(+) pumps (P2C and P2D ATPases) as well as a primitive H(+)-ATPase. Both Na(+) and H(+) pumps also co-exist in the moss P. patens, which has a P2D Na(+)-ATPase. In contrast to the primitive H(+)-ATPases in chlorophytes and P. patens, the H(+)-ATPases from vascular plants all have a large C-terminal regulatory domain as well as a conserved Arg in transmembrane segment 5 that is predicted to function as part of a backflow protection mechanism. Together these features are predicted to enable H(+) pumps in vascular plants to create large electrochemical gradients that can be modulated in response to diverse physiological cues. The complete inventory of P-type ATPases in the major branches of Viridiplantae is an important starting point for elucidating the evolution in plants of these important pumps.
Keywords: Na+ pumps; P-type ATPases; evolution; plants; salt tolerance.
Figures





References
LinkOut - more resources
Full Text Sources