

Lack of PPARγ in myeloid cells confers resistance to Listeria monocytogenes infection
- PMID: 22629382
- PMCID: PMC3357414
- DOI: 10.1371/journal.pone.0037349
Lack of PPARγ in myeloid cells confers resistance to Listeria monocytogenes infection
Abstract
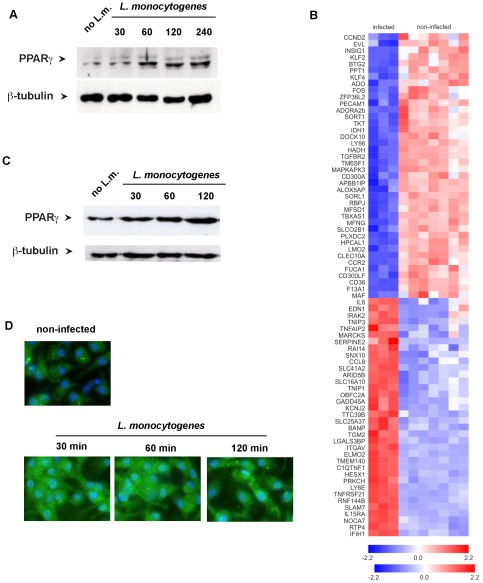
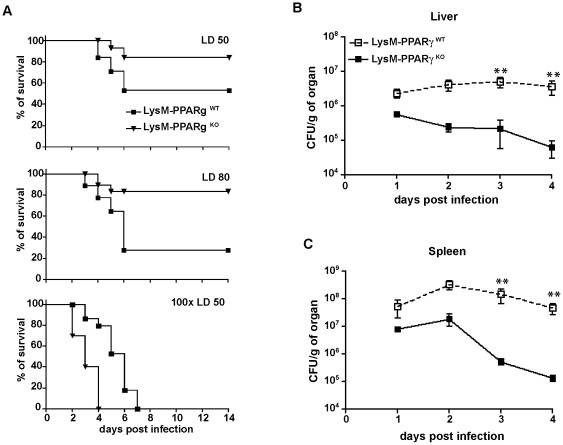
The peroxisomal proliferator-activated receptor γ (PPARγ) is a nuclear receptor that controls inflammation and immunity. Innate immune defense against bacterial infection appears to be compromised by PPARγ. The relevance of PPARγ in myeloid cells, that organize anti-bacterial immunity, for the outcome of immune responses against intracellular bacteria such as Listeria monocytogenes in vivo is unknown. We found that Listeria monocytogenes infection of macrophages rapidly led to increased expression of PPARγ. This prompted us to investigate whether PPARγ in myeloid cells influences innate immunity against Listeria monocytogenes infection by using transgenic mice with myeloid-cell specific ablation of PPARγ (LysMCre×PPARγ(flox/flox)). Loss of PPARγ in myeloid cells results in enhanced innate immune defense against Listeria monocytogenes infection both, in vitro and in vivo. This increased resistance against infection was characterized by augmented levels of bactericidal factors and inflammatory cytokines: ROS, NO, IFNγ TNF IL-6 and IL-12. Moreover, myeloid cell-specific loss of PPARγ enhanced chemokine and adhesion molecule expression leading to improved recruitment of inflammatory Ly6C(hi) monocytes to sites of infection. Importantly, increased resistance against Listeria infection in the absence of PPARγ was not accompanied by enhanced immunopathology. Our results elucidate a yet unknown regulatory network in myeloid cells that is governed by PPARγ and restrains both listeriocidal activity and recruitment of inflammatory monocytes during Listeria infection, which may contribute to bacterial immune escape. Pharmacological interference with PPARγ activity in myeloid cells might represent a novel strategy to overcome intracellular bacterial infection.
Conflict of interest statement
Figures





References
-
- Glass CK, Saijo K. Nuclear receptor transrepression pathways that regulate inflammation in macrophages and T cells. Nature reviews Immunology. 2010;10:365–376. - PubMed
-
- Marx N, Sukhova G, Murphy C, Libby P, Plutzky J. Macrophages in human atheroma contain PPARgamma: differentiation-dependent peroxisomal proliferator-activated receptor gamma(PPARgamma) expression and reduction of MMP-9 activity through PPARgamma activation in mononuclear phagocytes in vitro. Am J Pathol. 1998;153:17–23. - PMC - PubMed
-
- Clark RB, Bishop-Bailey D, Estrada-Hernandez T, Hla T, Puddington L, et al. The nuclear receptor PPAR gamma and immunoregulation: PPAR gamma mediates inhibition of helper T cell responses. J Immunol. 2000;164:1364–1371. - PubMed
-
- Jiang C, Ting AT, Seed B. PPAR-gamma agonists inhibit production of monocyte inflammatory cytokines. Nature. 1998;391:82–86. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
