

Review
doi: 10.1016/j.neuron.2012.04.023.
How we feel: ion channel partnerships that detect mechanical inputs and give rise to touch and pain perception
Affiliations
- PMID: 22632719
- PMCID: PMC3371091
- DOI: 10.1016/j.neuron.2012.04.023
Item in Clipboard
Review
How we feel: ion channel partnerships that detect mechanical inputs and give rise to touch and pain perception
Neuron.
.
Abstract
Every moment of every day, our skin and its embedded sensory neurons are bombarded with mechanical cues that we experience as pleasant or painful. Knowing the difference between innocuous and noxious mechanical stimuli is critical for survival and relies on the function of mechanoreceptor neurons that vary in their size, shape, and sensitivity. Their function is poorly understood at the molecular level. This review emphasizes the importance of integrating analysis at the molecular and cellular levels and focuses on the discovery of ion channel proteins coexpressed in the mechanoreceptors of worms, flies, and mice.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
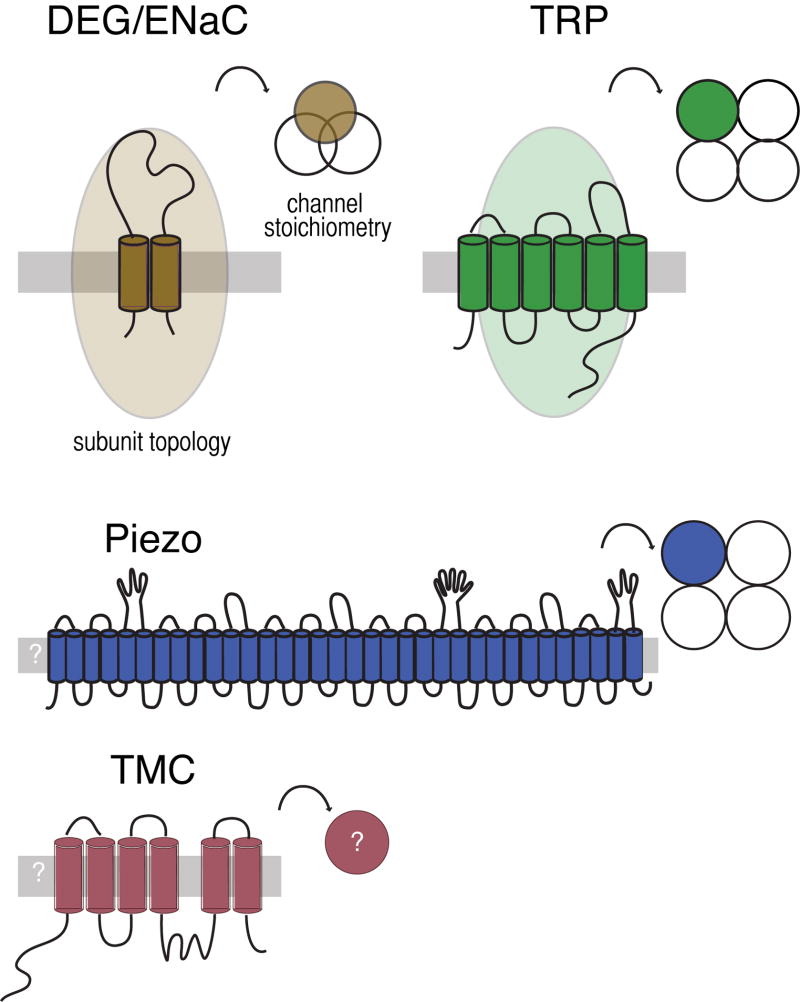
The TRP channel genes are conserved in eukaryotes and encode proteins predicted to have six transmembrane domains and assemble into tetrameric ion channels. Many TRPs have ankyrin repeats in their intracellular amino terminal; some have more than ten such repeats (Venkatachalam and Montell, 2007). The DEG/ENaC genes are absent from plants, yeast and other microbes, but conserved in animals (Goodman and Schwarz, 2003). They encode proteins with two transmembrane domains and a large extracellular domain. Three DEG/ENaC proteins assemble to form an ion channel. Both TRP channels and DEG/ENaC proteins can form homomeric and heteromeric channels, increasing the potential for channel diversity. Recently, two additional classes of membrane proteins (Piezo and TMC) have been linked to mechanotransduction in mammals (Coste et al, 2010; Kawashima et al, 2011) and Drosophila fruit flies (Kim et al., 2012). Piezo is sufficient to produce stretch activated channels in heterologous cells (Coste et al, 2010; Coste et al., 2012; Bae et al., 2011) and purified Piezo forms a channel in lipid bilayers (Coste et al., 2012). TMC1 and TMC2 are required for mechanotransduction by sound- and vibration-sensing hair cells in mice (Kawashima et al, 2011). Both Piezo and TMC have homologs in invertebrates; the C. elegans TMC homolog is expressed in the multidendritic PVD nociceptors. The predicted topology and stoichiometry of Piezo and TMC await further experimental confirmation.
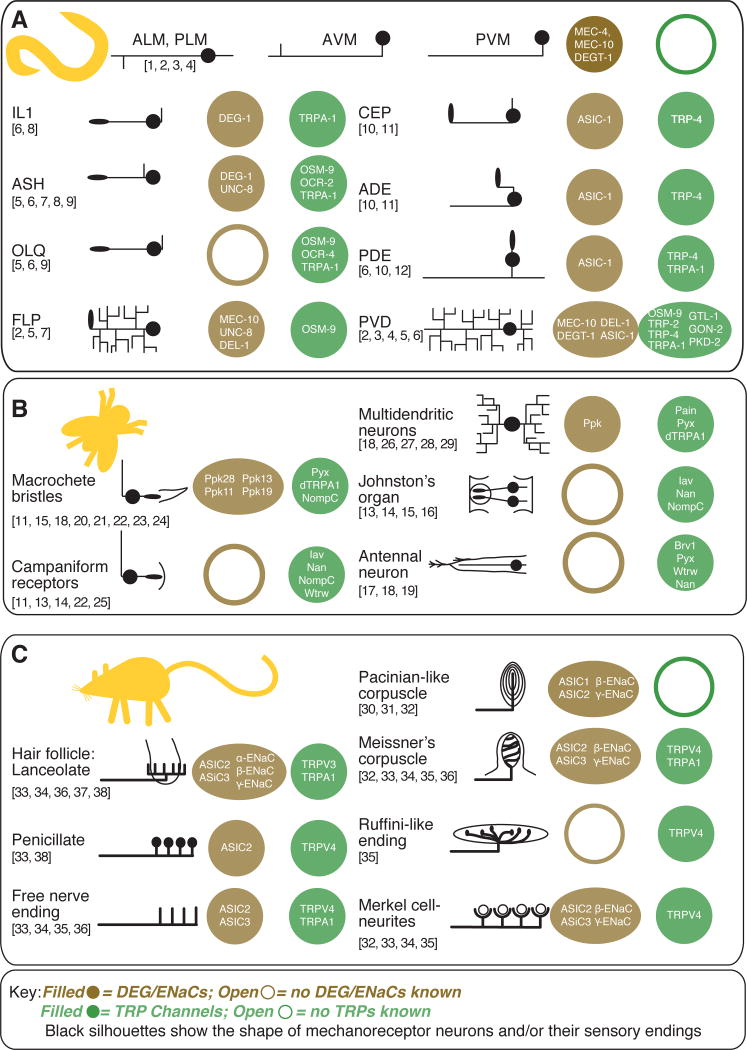
This graphical table illustrates the gross morphology of entire mechanoreceptor neurons (C. elegans) or peripheral sensory endings (Drosophila, mice) and lists ion channel subunits that are expressed in each class of mechanoreceptor cell. Sources for C. elegans mechanoreceptor expression are listed by number above: 1) Driscoll and Chalfie, 1991, 2) Huang and Chalfie, 1994, 3) Chatzigeorgiou et al., 2010, 4) Smith et al., 2010, 5) Colbert et al., 1997, 6) Kindt et al., 2007, 7) Tavernarakis et al., 1997, 8) Hall et al., 1997, 9) Tobin et al., 2002, 10) Voglis and Tavernarakis, 2008, 11) Walker et al., 2000, 12) Li et al., 2006. Drosophila melanogaster sourcesare: 13) Gong et al., 2004, 14) Kim et al., 2003, 15) Lee et al., 2010, 16) Liang et al., 2011, 17) Gallio et al., 2011, 18) Lee et al., 2005, 19) Liu et al., 2007, 20) Hamada et al., 2008, 21) Kim et al., 2010, 22) Cheng et al., 2010, 23) Chen et al., 2010, 24) Liu et al., 2003, 25) Bechstedt et al., 2010, 26) Tracey et al., 2003, 27) Zhong et al., 2012, 28) Adams et al., 1998, 29) Zhong et al., 2010). These sources establishing expression in peripheral endings in mice and humans were consulted: 30) Calavia et al., 2010, 31) Montaño et al., 2009, 32)Drummond et al., 2000, 33) García-Añoveros et al., 2001, 34) Price et al., 2001, 35) Suzuki et al., 2003b, 36) Kwan et al., 2009, 37) Price et al., 2000, 38) Fricke et al., 2000, 39) Xu et al., 2002).
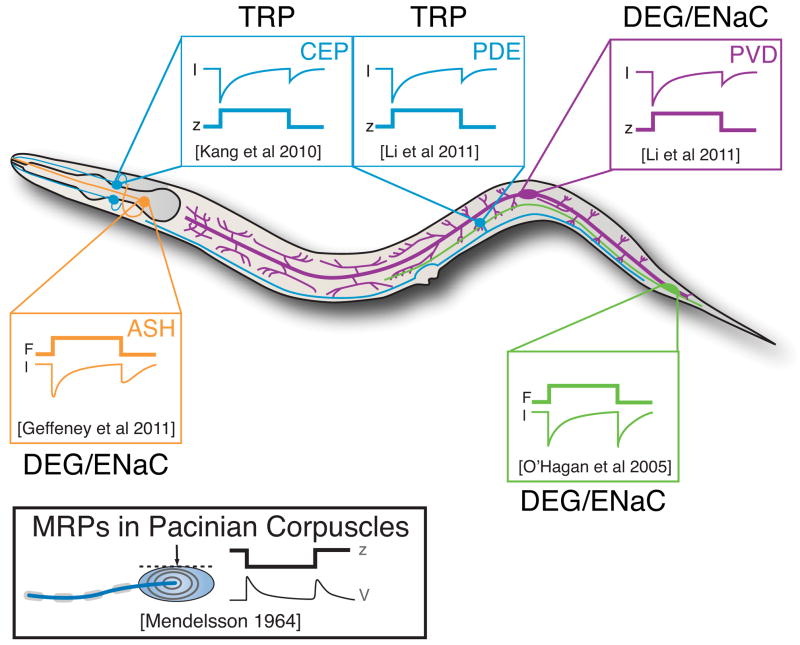
Mechanoreceptor currents have been recorded using in vivo whole-cell patch clamp recording and the predominant ion channel type identified by genetic dissection. Traces adapted from the following: PLM: O’Hagan et al. (2005); CEP: Kang et al. (2010); PDE, PVD: Li et al. (2011b); ASH: Geffeney et al. (2011). Displacement stimuli were applied to activate CEP, PDE, and PVD, while mechanical stimuli delivering known forces were applied to activate PLM and ASH. Receptor potentials in PLM and ASH mirror the receptor currents (Geffeney et al., 2011; O’Hagan et al., 2005) and are reminiscent of the response dynamics of Pacinian corpuscles in cats (inset, lower left).
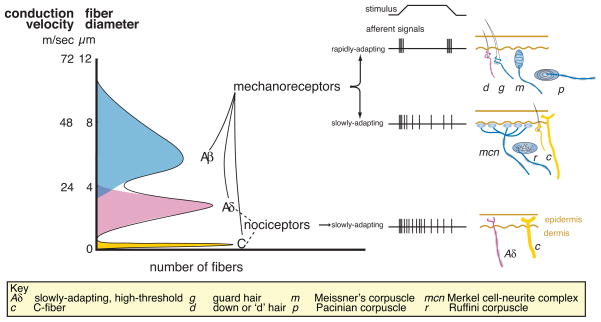
Mammalian mechanoreceptor nerve fibers are classified by according to three physiological properties: 1) the speed of action potential propagation (which depends on fiber diameter and myelination state); 2) the threshold for activation; and 3) the rate of adaptation to mechanical stimuli. The broad categories of Aβ, Aδ and C-fibers are defined by their propagation speeds where Aβ-fibers have the most rapid propagation speeds and the slender, unmyelinated C-fibers have the slowest. Most fibers in these categories share other properties. For example, C fibers have slow rates of adaptation to mechanical stimuli and many have high mechanical thresholds. In contrast, most Aβ-fibers have low mechanical thresholds and these fibers are thought to innervate light-touch receptors in the skin. The challenge is to link fiber properties to the diverse endings in the skin.
Similar articles
-
Piezo2 voltage-block regulates mechanical pain sensitivity.Brain. 2024 Oct 3;147(10):3487-3500. doi: 10.1093/brain/awae227. Brain. 2024. PMID: 38984717 Free PMC article.
-
TRPC1 contributes to light-touch sensation and mechanical responses in low-threshold cutaneous sensory neurons.J Neurophysiol. 2012 Feb;107(3):913-22. doi: 10.1152/jn.00658.2011. Epub 2011 Nov 9. J Neurophysiol. 2012. PMID: 22072513 Free PMC article.
-
PKCγ interneurons, a gateway to pathological pain in the dorsal horn.J Neural Transm (Vienna). 2020 Apr;127(4):527-540. doi: 10.1007/s00702-020-02162-6. Epub 2020 Feb 27. J Neural Transm (Vienna). 2020. PMID: 32108249 Review.
-
The DRASIC cation channel contributes to the detection of cutaneous touch and acid stimuli in mice.Neuron. 2001 Dec 20;32(6):1071-83. doi: 10.1016/s0896-6273(01)00547-5. Neuron. 2001. PMID: 11754838
-
Transduction and encoding sensory information by skin mechanoreceptors.Pflugers Arch. 2015 Jan;467(1):109-19. doi: 10.1007/s00424-014-1651-7. Epub 2014 Nov 23. Pflugers Arch. 2015. PMID: 25416542 Review.
Cited by
-
Runx1 controls terminal morphology and mechanosensitivity of VGLUT3-expressing C-mechanoreceptors.J Neurosci. 2013 Jan 16;33(3):870-82. doi: 10.1523/JNEUROSCI.3942-12.2013. J Neurosci. 2013. PMID: 23325226 Free PMC article.
-
Insight into DEG/ENaC channel gating from genetics and structure.Physiology (Bethesda). 2012 Oct;27(5):282-90. doi: 10.1152/physiol.00006.2012. Physiology (Bethesda). 2012. PMID: 23026751 Free PMC article. Review.
-
A TRP channel trio mediates acute noxious heat sensing.Nature. 2018 Mar 29;555(7698):662-666. doi: 10.1038/nature26137. Epub 2018 Mar 14. Nature. 2018. PMID: 29539642
-
Stomatin-domain protein interactions with acid-sensing ion channels modulate nociceptor mechanosensitivity.J Physiol. 2013 Nov 15;591(22):5555-74. doi: 10.1113/jphysiol.2013.261180. Epub 2013 Aug 19. J Physiol. 2013. PMID: 23959680 Free PMC article.
-
The topographical attenuation of cutaneous input is modulated at the ankle joint during gait.Exp Brain Res. 2024 Jan;242(1):149-161. doi: 10.1007/s00221-023-06737-z. Epub 2023 Nov 18. Exp Brain Res. 2024. PMID: 37979067
References
-
- Akopian AN, Chen CC, Ding Y, Cesare P, Wood JN. A new member of the acid-sensing ion channel family. Neuroreport. 2000;11:2217–2222. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
