

Balance of activity between LN(v)s and glutamatergic dorsal clock neurons promotes robust circadian rhythms in Drosophila
- PMID: 22632728
- PMCID: PMC3361687
- DOI: 10.1016/j.neuron.2012.02.034
Balance of activity between LN(v)s and glutamatergic dorsal clock neurons promotes robust circadian rhythms in Drosophila
Erratum in
- Neuron. 2012 Jun 21;74(6):1138
Abstract
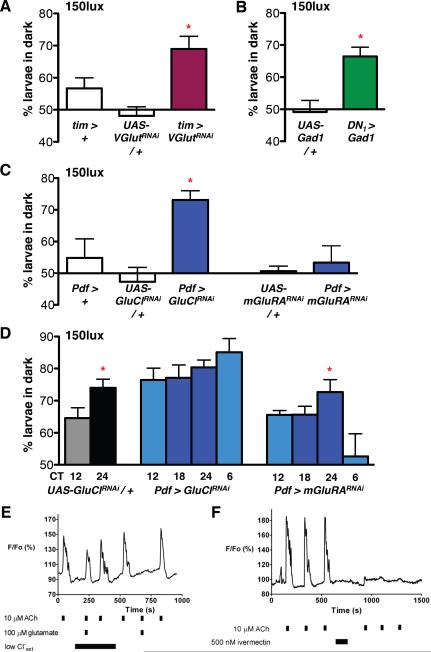
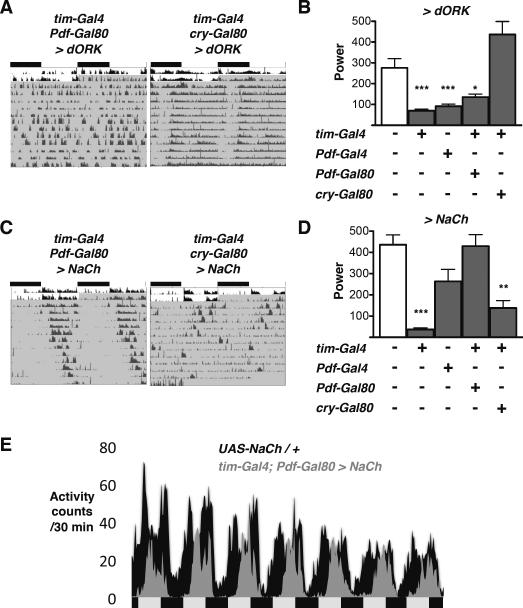
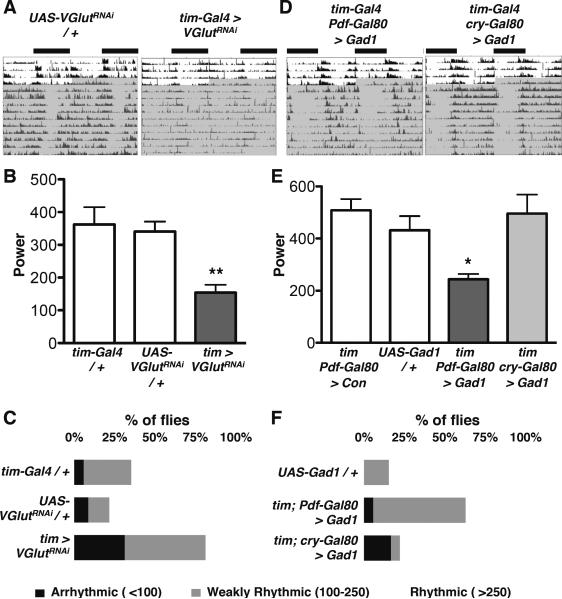
Circadian rhythms offer an excellent opportunity to dissect the neural circuits underlying innate behavior because the genes and neurons involved are relatively well understood. We first sought to understand how Drosophila clock neurons interact in the simple circuit that generates circadian rhythms in larval light avoidance. We used genetics to manipulate two groups of clock neurons, increasing or reducing excitability, stopping their molecular clocks, and blocking neurotransmitter release and reception. Our results revealed that lateral neurons (LN(v)s) promote and dorsal clock neurons (DN(1)s) inhibit light avoidance, these neurons probably signal at different times of day, and both signals are required for rhythmic behavior. We found that similar principles apply in the more complex adult circadian circuit that generates locomotor rhythms. Thus, the changing balance in activity between clock neurons with opposing behavioral effects generates robust circadian behavior and probably helps organisms transition between discrete behavioral states, such as sleep and wakefulness.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
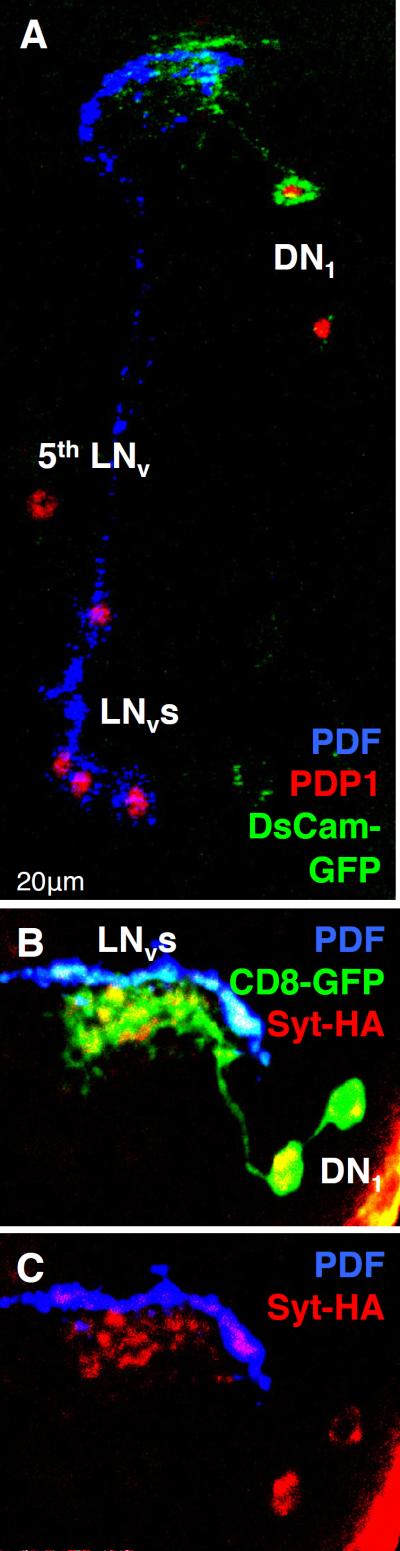
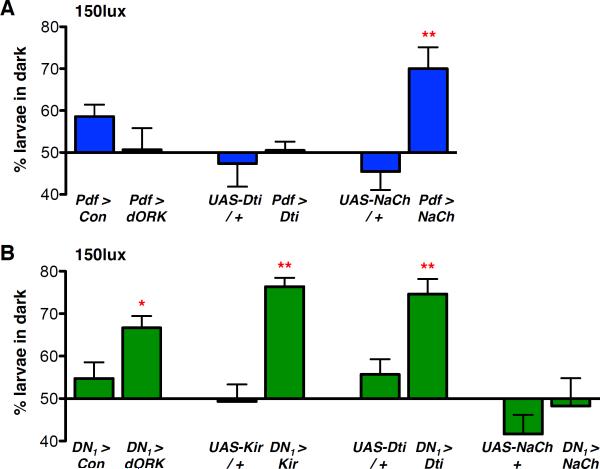
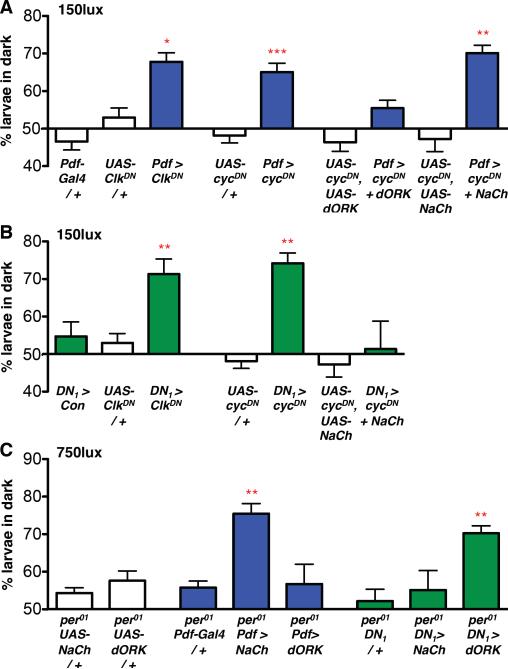
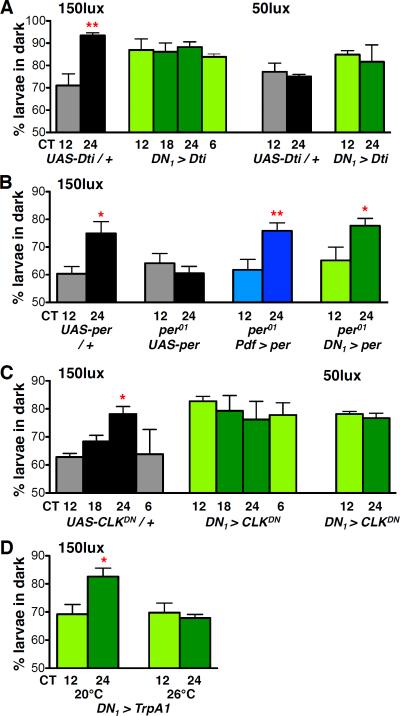
References
-
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
-
- Cully DF, Vassilatis DK, Liu KK, Paress PS, Van der Ploeg LH, Schaeffer JM, Arena JP. Cloning of an avermectin-sensitive glutamate-gated chloride channel from Caenorhabditis elegans. Nature. 1994;371:707–711. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
