

Understanding the development of roots exposed to contaminants and the potential of plant-associated bacteria for optimization of growth
- PMID: 22634257
- PMCID: PMC3394651
- DOI: 10.1093/aob/mcs105
Understanding the development of roots exposed to contaminants and the potential of plant-associated bacteria for optimization of growth
Abstract
Background and scope: Plant responses to the toxic effects of soil contaminants, such as excess metals or organic substances, have been studied mainly at physiological, biochemical and molecular levels, but the influence on root system architecture has received little attention. Nevertheless, the precise position, morphology and extent of roots can influence contaminant uptake. Here, data are discussed that aim to increase the molecular and ecological understanding of the influence of contaminants on root system architecture. Furthermore, the potential of plant-associated bacteria to influence root growth by their growth-promoting and stress-relieving capacities is explored.
Methods: Root growth parameters of Arabidopsis thaliana seedlings grown in vertical agar plates are quantified. Mutants are used in a reverse genetics approach to identify molecular components underlying quantitative changes in root architecture after exposure to excess cadmium, copper or zinc. Plant-associated bacteria are isolated from contaminated environments, genotypically and phenotypically characterized, and used to test plant root growth improvement in the presence of contaminants.
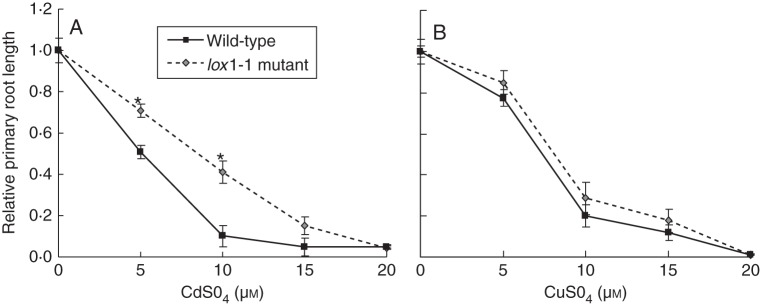
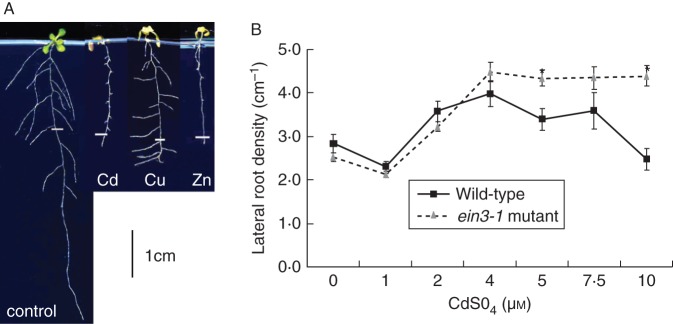
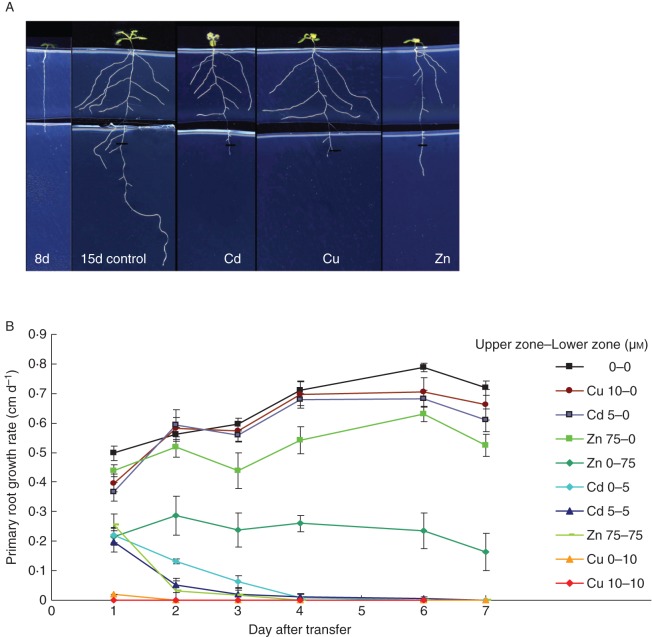
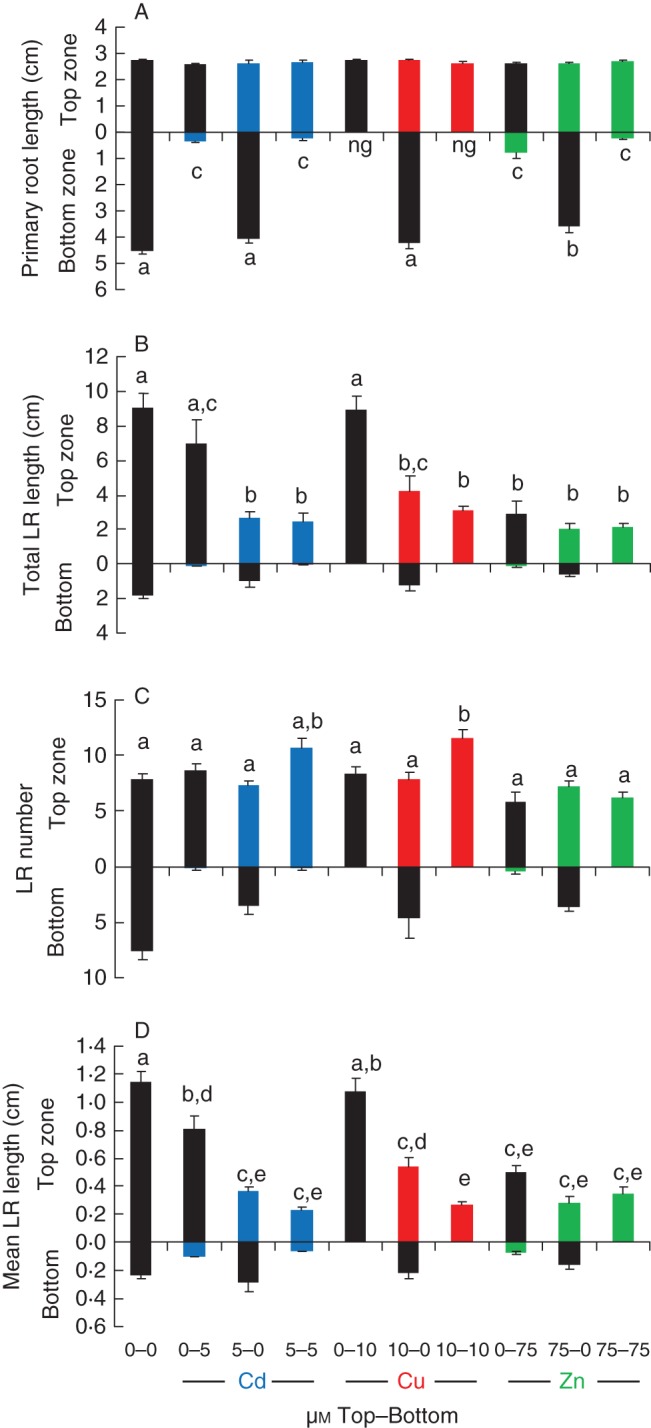
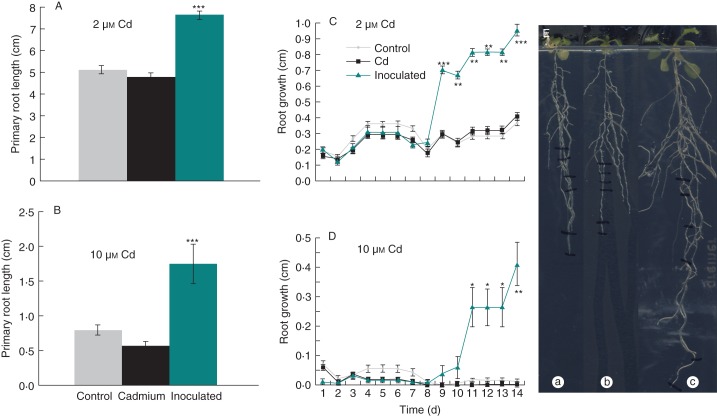
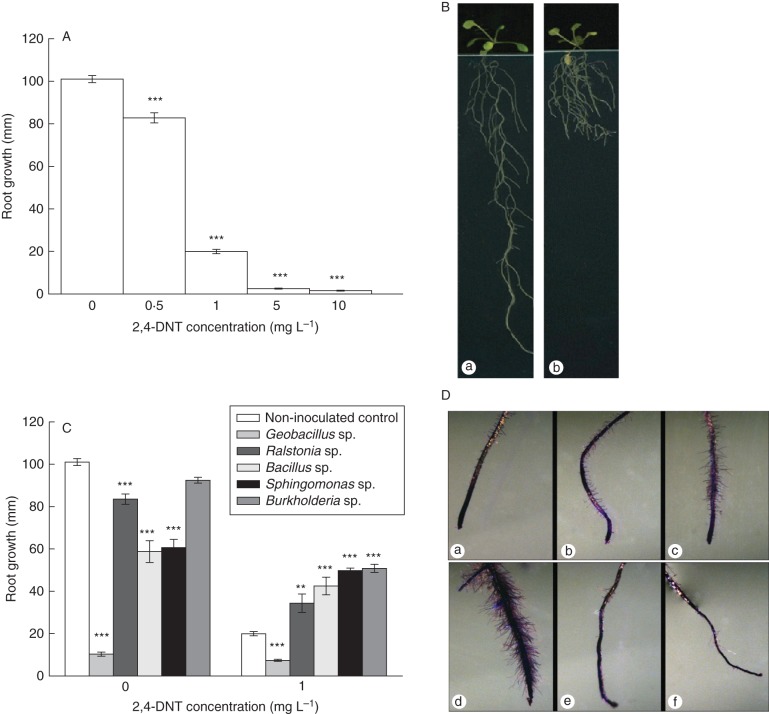
Key results: The molecular determinants of primary root growth inhibition and effects on lateral root density by cadmium were identified. A vertical split-root system revealed local effects of cadmium and copper on root development. However, systemic effects of zinc exposure on root growth reduced both the avoidance of contaminated areas and colonization of non-contaminated areas. The potential for growth promotion and contaminant degradation of plant-associated bacteria was demonstrated by improved root growth of inoculated plants exposed to 2,4-di-nitro-toluene (DNT) or cadmium.
Conclusions: Knowledge concerning the specific influence of different contaminants on root system architecture and the molecular mechanisms by which this is achieved can be combined with the exploitation of plant-associated bacteria to influence root development and increase plant stress tolerance, which should lead to more optimal root systems for application in phytoremediation or safer biomass production.
Figures
Similar articles
-
Root architecture and morphometric analysis of Arabidopsis thaliana grown in Cd/Cu/Zn-gradient agar dishes: A new screening technique for studying plant response to metals.Plant Physiol Biochem. 2015 Jun;91:20-7. doi: 10.1016/j.plaphy.2015.03.010. Epub 2015 Mar 28. Plant Physiol Biochem. 2015. PMID: 25839424
-
Plant architecture, auxin homeostasis and phenol content in Arabidopsis thaliana grown in cadmium- and zinc-enriched media.J Plant Physiol. 2017 Sep;216:174-180. doi: 10.1016/j.jplph.2017.06.008. Epub 2017 Jul 3. J Plant Physiol. 2017. PMID: 28704702
-
Response to copper excess in Arabidopsis thaliana: Impact on the root system architecture, hormone distribution, lignin accumulation and mineral profile.Plant Physiol Biochem. 2010 Aug;48(8):673-82. doi: 10.1016/j.plaphy.2010.05.005. Epub 2010 May 27. Plant Physiol Biochem. 2010. PMID: 20542443
-
Plant responses to abiotic stresses: heavy metal-induced oxidative stress and protection by mycorrhization.J Exp Bot. 2002 May;53(372):1351-65. J Exp Bot. 2002. PMID: 11997381 Review.
-
Arabidopsis root growth and development under metal exposure presented in an adverse outcome pathway framework.Plant Cell Environ. 2022 Mar;45(3):737-750. doi: 10.1111/pce.14147. Epub 2021 Jul 17. Plant Cell Environ. 2022. PMID: 34240430 Free PMC article. Review.
Cited by
-
Genetic Circuit Design in Rhizobacteria.Biodes Res. 2022 Sep 1;2022:9858049. doi: 10.34133/2022/9858049. eCollection 2022. Biodes Res. 2022. PMID: 37850138 Free PMC article. Review.
-
Cadmium and Plant Development: An Agony from Seed to Seed.Int J Mol Sci. 2019 Aug 15;20(16):3971. doi: 10.3390/ijms20163971. Int J Mol Sci. 2019. PMID: 31443183 Free PMC article. Review.
-
An automated, cost-effective and scalable, flood-and-drain based root phenotyping system for cereals.Plant Methods. 2016 Jun 24;12:34. doi: 10.1186/s13007-016-0135-5. eCollection 2016. Plant Methods. 2016. PMID: 27347000 Free PMC article.
-
Toxicity responses of Cu and Cd: the involvement of miRNAs and the transcription factor SPL7.BMC Plant Biol. 2016 Jun 28;16(1):145. doi: 10.1186/s12870-016-0830-4. BMC Plant Biol. 2016. PMID: 27352843 Free PMC article.
-
Nicotiana tabacum-associated bioengineered Pseudomonas putida can enhance rhizoremediation of soil containing 2,4-dinitrotoluene.3 Biotech. 2020 Sep;10(9):398. doi: 10.1007/s13205-020-02395-y. Epub 2020 Aug 19. 3 Biotech. 2020. PMID: 32864284 Free PMC article.
References
-
- Alford E, Pilon-Smits E, Paschke M. Metallophytes – a view from the rhizosphere. Plant and Soil. 2010;337:33–50.
-
- Armengaud P, Zambaux K, Hills A, et al. EZ-Rhizo: integrated software for the fast and accurate measurement of root system architecture. Plant Journal. 2009;57:945–956. - PubMed
-
- Bardos P, Chapman T, Andersson-Sköld Y, et al. Biomass production on marginal land. Biocycle. 2008;49:50–52.
-
- Baluška F, Mancuso S, Volkmann D, Barlow P. Root apex transition zone: a signalling-response nexus in the root. Trends in Plant Science. 2010;15:402–408. - PubMed
-
- Belimov AA, Hontzeas N, Safronova VI, et al. Cadmium-tolerant plant growth-promoting bacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.) Soil Biology & Biochemistry. 2005;37:241–250.
