

RNA 3'-end mismatch excision by the severe acute respiratory syndrome coronavirus nonstructural protein nsp10/nsp14 exoribonuclease complex
- PMID: 22635272
- PMCID: PMC3386072
- DOI: 10.1073/pnas.1201130109
RNA 3'-end mismatch excision by the severe acute respiratory syndrome coronavirus nonstructural protein nsp10/nsp14 exoribonuclease complex
Abstract
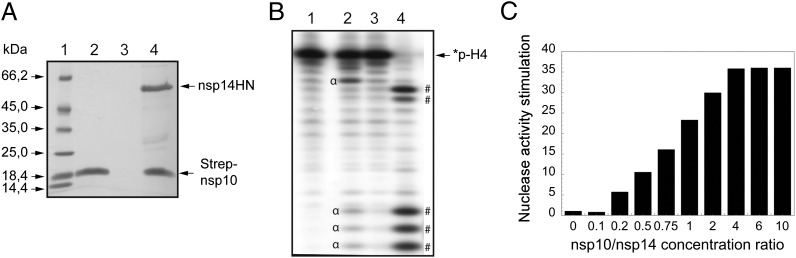
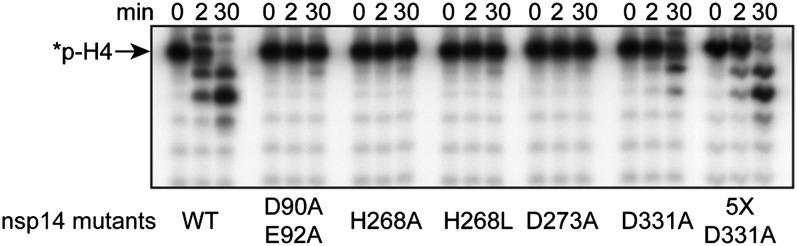
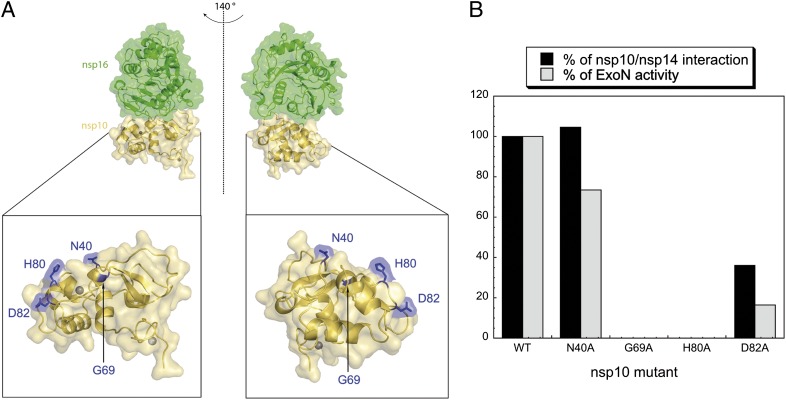
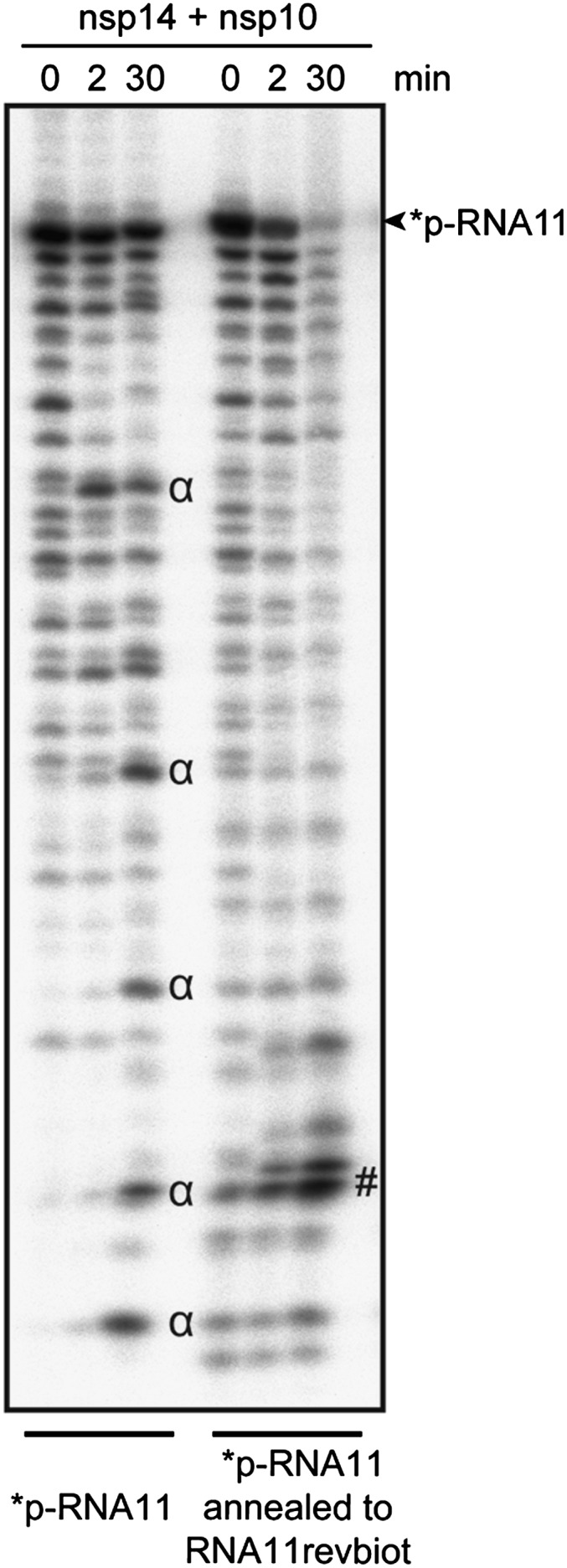
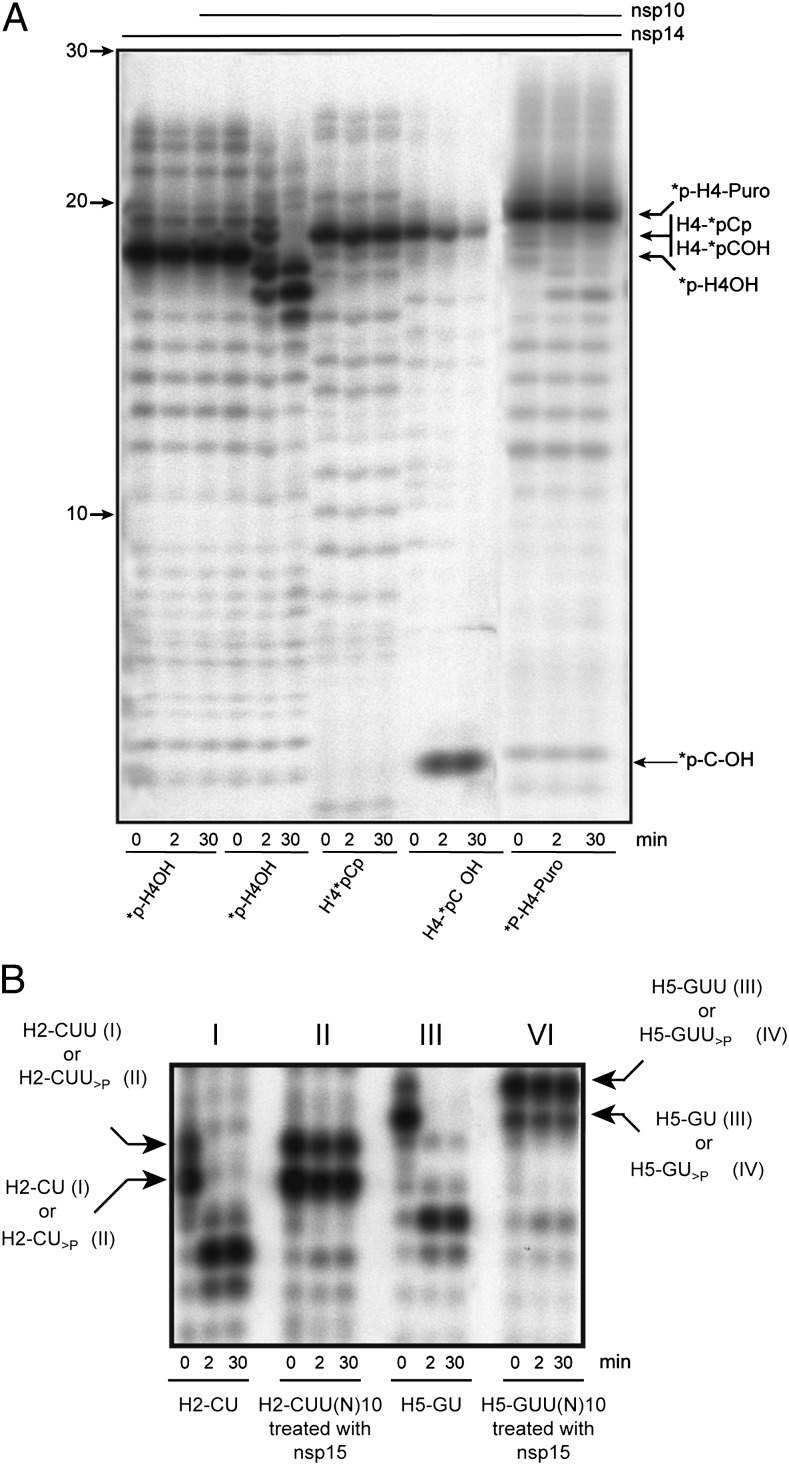
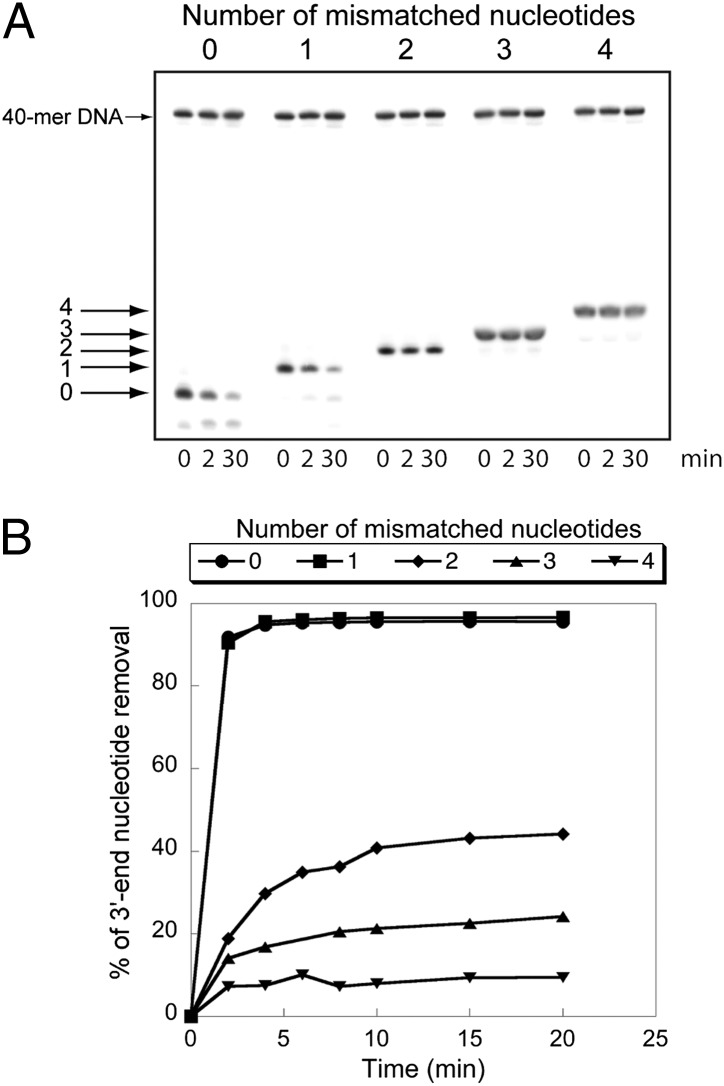
The replication/transcription complex of severe acute respiratory syndrome coronavirus is composed of at least 16 nonstructural proteins (nsp1-16) encoded by the ORF-1a/1b. This complex includes replication enzymes commonly found in positive-strand RNA viruses, but also a set of RNA-processing activities unique to some nidoviruses. The nsp14 protein carries both exoribonuclease (ExoN) and (guanine-N7)-methyltransferase (N7-MTase) activities. The nsp14 ExoN activity ensures a yet-uncharacterized function in the virus life cycle and must be regulated to avoid nonspecific RNA degradation. In this work, we show that the association of nsp10 with nsp14 stimulates >35-fold the ExoN activity of the latter while playing no effect on N7-MTase activity. Nsp10 mutants unable to interact with nsp14 are not proficient for ExoN activation. The nsp10/nsp14 complex hydrolyzes double-stranded RNA in a 3' to 5' direction as well as a single mismatched nucleotide at the 3'-end mimicking an erroneous replication product. In contrast, di-, tri-, and longer unpaired ribonucleotide stretches, as well as 3'-modified RNAs, resist nsp10/nsp14-mediated excision. In addition to the activation of nsp16-mediated 2'-O-MTase activity, nsp10 also activates nsp14 in an RNA processing function potentially connected to a replicative mismatch repair mechanism.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
