

Mechanistic insights into the retaining glucosyl-3-phosphoglycerate synthase from mycobacteria
- PMID: 22637481
- PMCID: PMC3397893
- DOI: 10.1074/jbc.M112.368191
Mechanistic insights into the retaining glucosyl-3-phosphoglycerate synthase from mycobacteria
Abstract
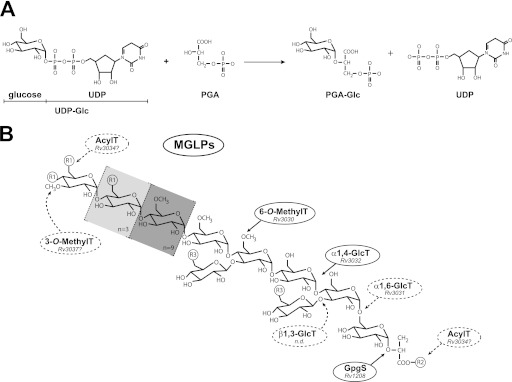
Considerable progress has been made in recent years in our understanding of the structural basis of glycosyl transfer. Yet the nature and relevance of the conformational changes associated with substrate recognition and catalysis remain poorly understood. We have focused on the glucosyl-3-phosphoglycerate synthase (GpgS), a "retaining" enzyme, that initiates the biosynthetic pathway of methylglucose lipopolysaccharides in mycobacteria. Evidence is provided that GpgS displays an unusually broad metal ion specificity for a GT-A enzyme, with Mg(2+), Mn(2+), Ca(2+), Co(2+), and Fe(2+) assisting catalysis. In the crystal structure of the apo-form of GpgS, we have observed that a flexible loop adopts a double conformation L(A) and L(I) in the active site of both monomers of the protein dimer. Notably, the L(A) loop geometry corresponds to an active conformation and is conserved in two other relevant states of the enzyme, namely the GpgS·metal·nucleotide sugar donor and the GpgS·metal·nucleotide·acceptor-bound complexes, indicating that GpgS is intrinsically in a catalytically active conformation. The crystal structure of GpgS in the presence of Mn(2+)·UDP·phosphoglyceric acid revealed an alternate conformation for the nucleotide sugar β-phosphate, which likely occurs upon sugar transfer. Structural, biochemical, and biophysical data point to a crucial role of the β-phosphate in donor and acceptor substrate binding and catalysis. Altogether, our experimental data suggest a model wherein the catalytic site is essentially preformed, with a few conformational changes of lateral chain residues as the protein proceeds along the catalytic cycle. This model of action may be applicable to a broad range of GT-A glycosyltransferases.
Figures





References
-
- Rini J., Esko J., Varki A. (2009) in Essentials of Glycobiology (Varki A., Cummings R., Esko J., Freeze H., Stanley P., Bertozzi C. R., Hart G., Etzler M. E., eds) 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY - PubMed
-
- Lairson L. L., Henrissat B., Davies G. J., Withers S. G. (2008) Glycosyltransferases. Structures, functions, and mechanisms. Annu. Rev. Biochem. 77, 521–555 - PubMed
-
- Persson K., Ly H. D., Dieckelmann M., Wakarchuk W. W., Withers S. G., Strynadka N. C. (2001) Crystal structure of the retaining galactosyltransferase LgtC from Neisseria meningitidis in complex with donor and acceptor sugar analogs. Nat. Struct. Biol. 8, 166–175 - PubMed
-
- Gibson R. P., Turkenburg J. P., Charnock S. J., Lloyd R., Davies G. J. (2002) Insights into trehalose synthesis provided by the structure of the retaining glucosyltransferase OtsA. Chem. Biol. 9, 1337–1346 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
