

Mutations in an Atypical TIR-NB-LRR-LIM Resistance Protein Confer Autoimmunity
- PMID: 22639607
- PMCID: PMC3355616
- DOI: 10.3389/fpls.2011.00071
Mutations in an Atypical TIR-NB-LRR-LIM Resistance Protein Confer Autoimmunity
Abstract
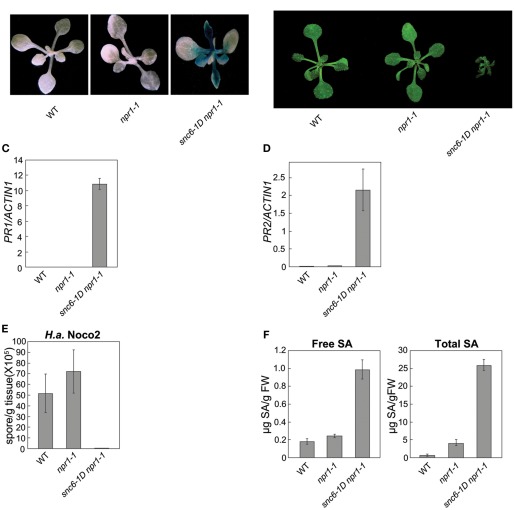
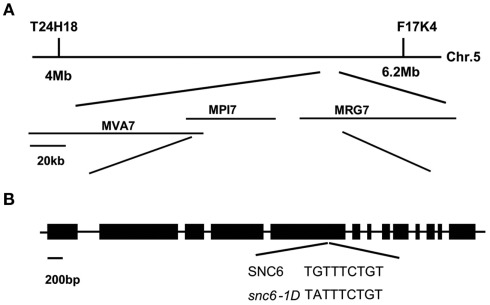
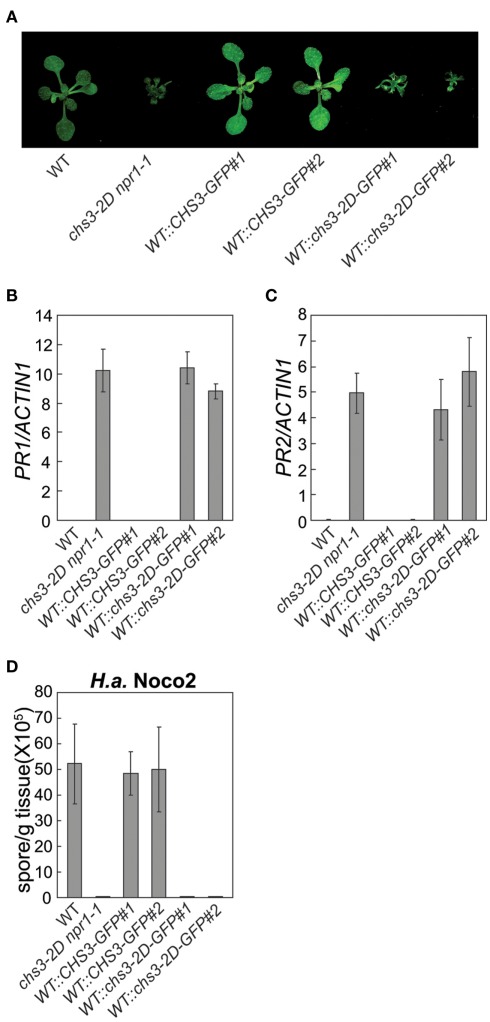
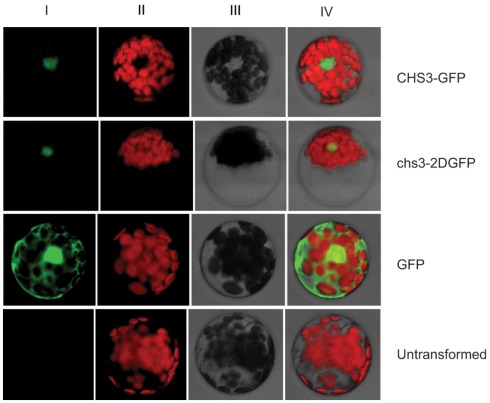
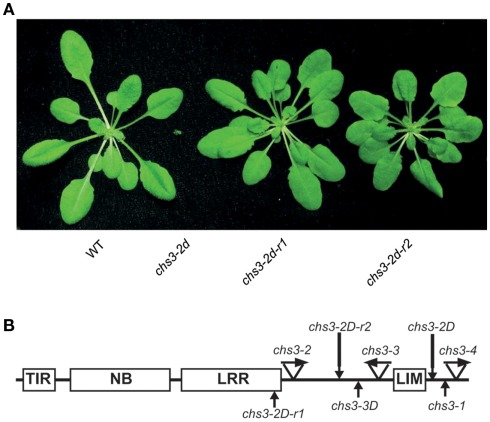
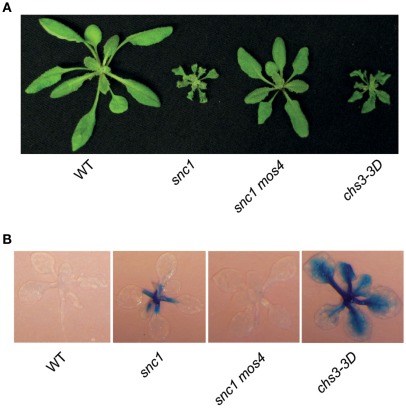
In order to defend against microbial infection, plants employ a complex immune system that relies partly on resistance (R) proteins that initiate intricate signaling cascades upon pathogen detection. The resistance signaling network utilized by plants is only partially characterized. A genetic screen conducted to identify novel defense regulators involved in this network resulted in the isolation of the snc6-1D mutant. Positional cloning revealed that this mutant contained a molecular lesion in the chilling sensitive 3 (CHS3) gene, thus the allele was renamed chs3-2D. CHS3 encodes a TIR-NB-LRR R protein that contains a C-terminal zinc-binding LIM (Lin-11, Isl-1, Mec-3) domain. Although this protein has been previously implicated in cold stress and defense response, the role of the LIM domain in modulating protein activity is unclear. The chs3-2D allele contains a G to A point mutation causing a C1340 to Y1340 substitution close to the LIM domain. It encodes a dominant gain-of-function mutation. The chs3-2D mutant is severely stunted and displays curled leaf morphology. Additionally, it constitutively expresses PATHOGENESIS-RELATED (PR) genes, accumulates salicylic acid, and shows enhanced resistance to the virulent oomycete isolate Hyaloperonospora arabidopsidis (H.a.) Noco2. Subcellular localization assays using GFP fusion constructs indicate that both CHS3 and chs3-2D localize to the nucleus. A third chs3 mutant allele, chs3-3D, was identified in an unrelated genetic screen in our lab. This allele contains a C to T point mutation resulting in an M1017 to V1017 substitution in the LRR-LIM linker region. Additionally, a chs3-2D suppressor screen identified two revertant alleles containing secondary mutations that abolish the mutant morphology. Analysis of the locations of these molecular lesions provides support for the hypothesis that the LIM domain represses CHS3 R-like protein activity. This repression may occur through either autoinhibition or binding of a negative defense regulator.
Keywords: Arabidopsis; CHS3; LIM domain; innate immunity; resistance protein.
Figures
References
-
- Aarts N., Metz M., Holub E., Staskawicz B. J., Daniels M. J., Parker J. E. (1998). Different requirements for EDS1 and NDR1 by disease resistance genes define at least two R gene-mediated signaling pathways in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 95, 10306–10311 10.1073/pnas.95.17.10306 - DOI - PMC - PubMed
-
- Baltz R., Evrard J. L., Bourdon V., Steinmetz A. (1996). The pollen-specific LIM protein PLIM-1 from sunflower binds nucleic acids in vitro. Sex. Plant Reprod. 9, 264–268 10.1007/BF02152700 - DOI
-
- Bernoux M., Ve T., Williams S., Warren C., Hatters D., Valkov E., Zhang X., Ellis J. G., Kobe B., Dodds P. N. (2011). Structural and functional analysis of a plant resistance protein TIR domain reveals interfaces for self-association, signaling, and autoregulation. Cell Host Microbe 9, 200–211 10.1016/j.chom.2011.02.009 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous