

Insulin analogs for the treatment of diabetes mellitus: therapeutic applications of protein engineering
- PMID: 22641195
- PMCID: PMC3360579
- DOI: 10.1111/j.1749-6632.2012.06468.x
Insulin analogs for the treatment of diabetes mellitus: therapeutic applications of protein engineering
Abstract
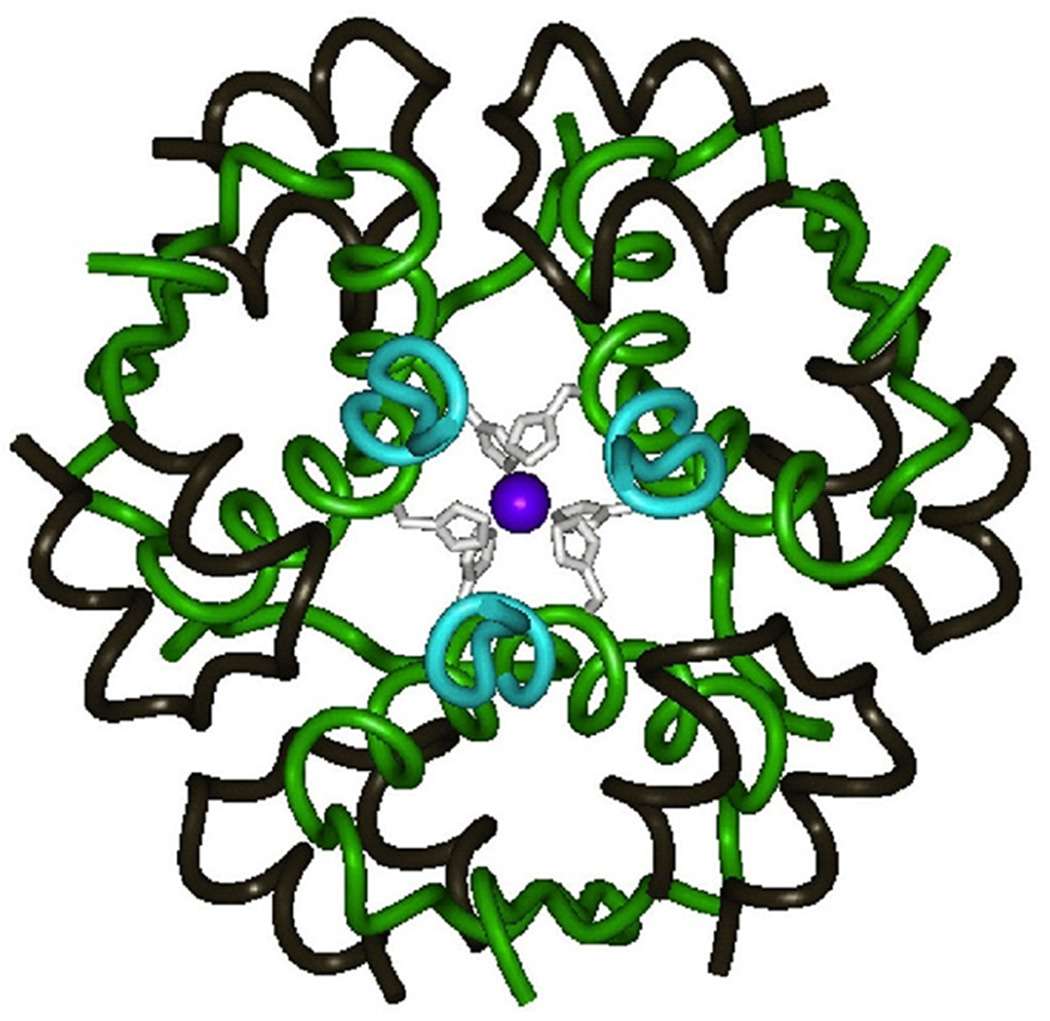
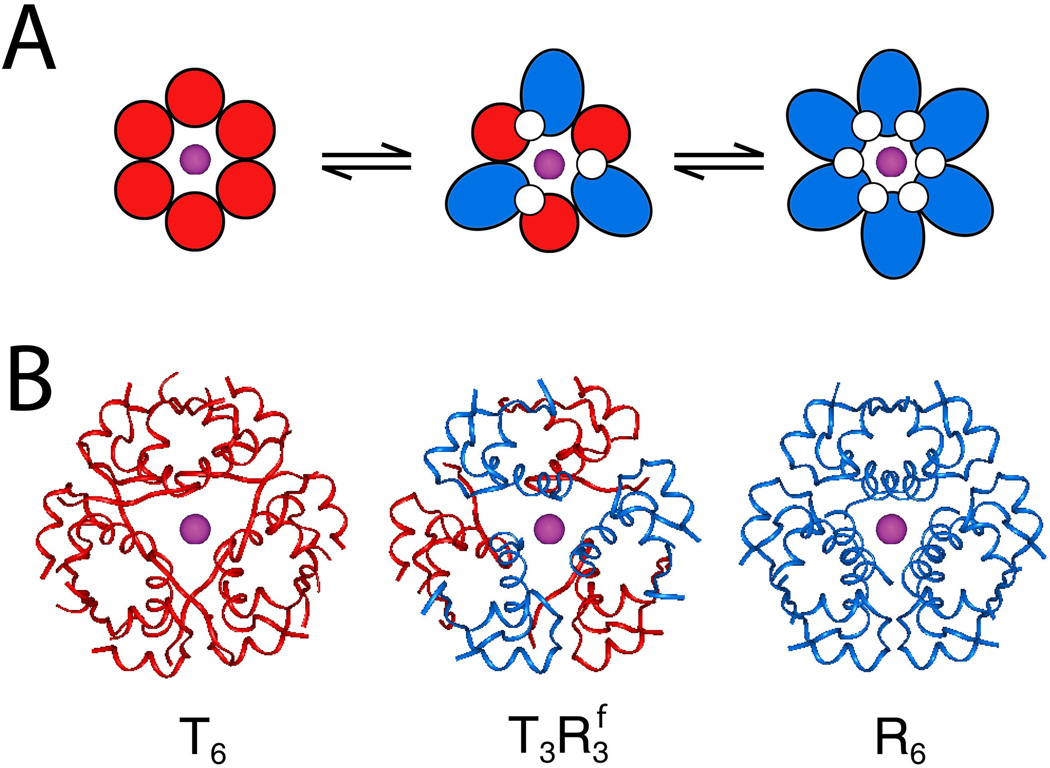
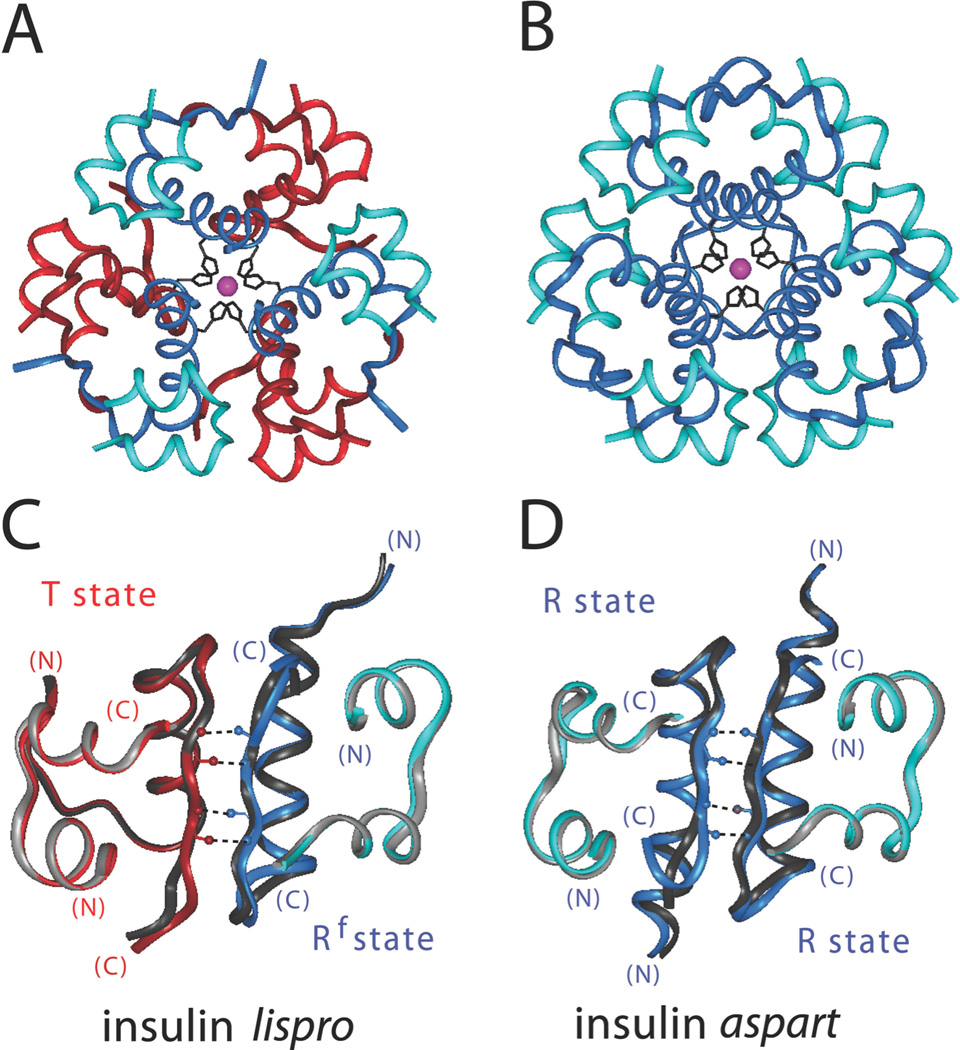
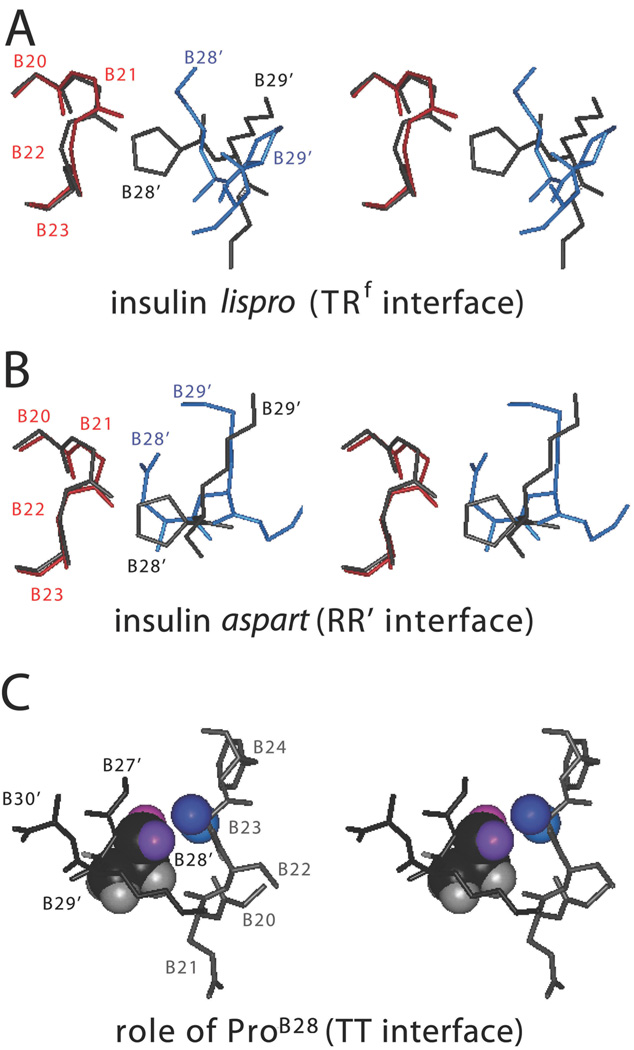
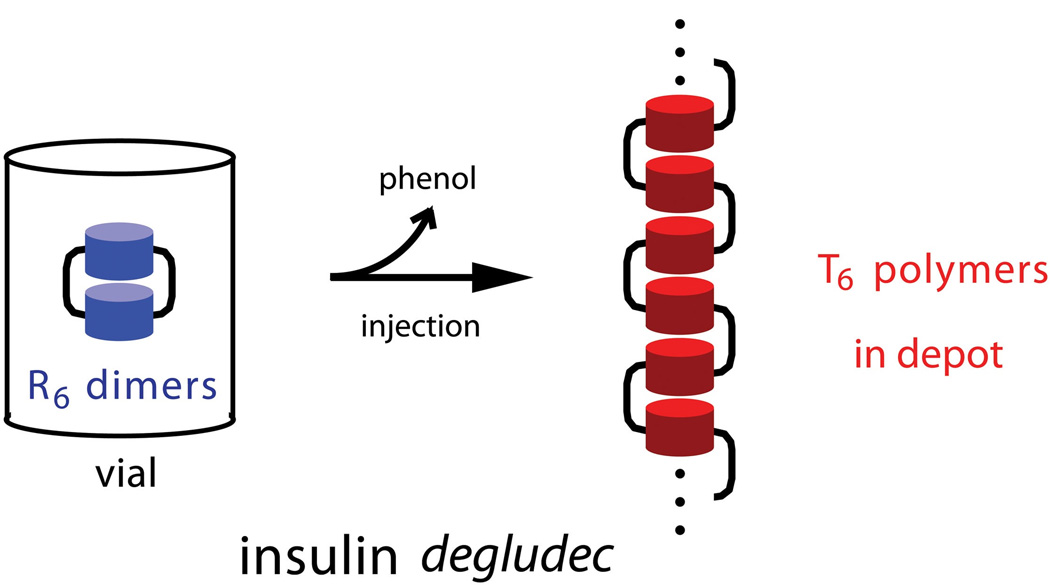
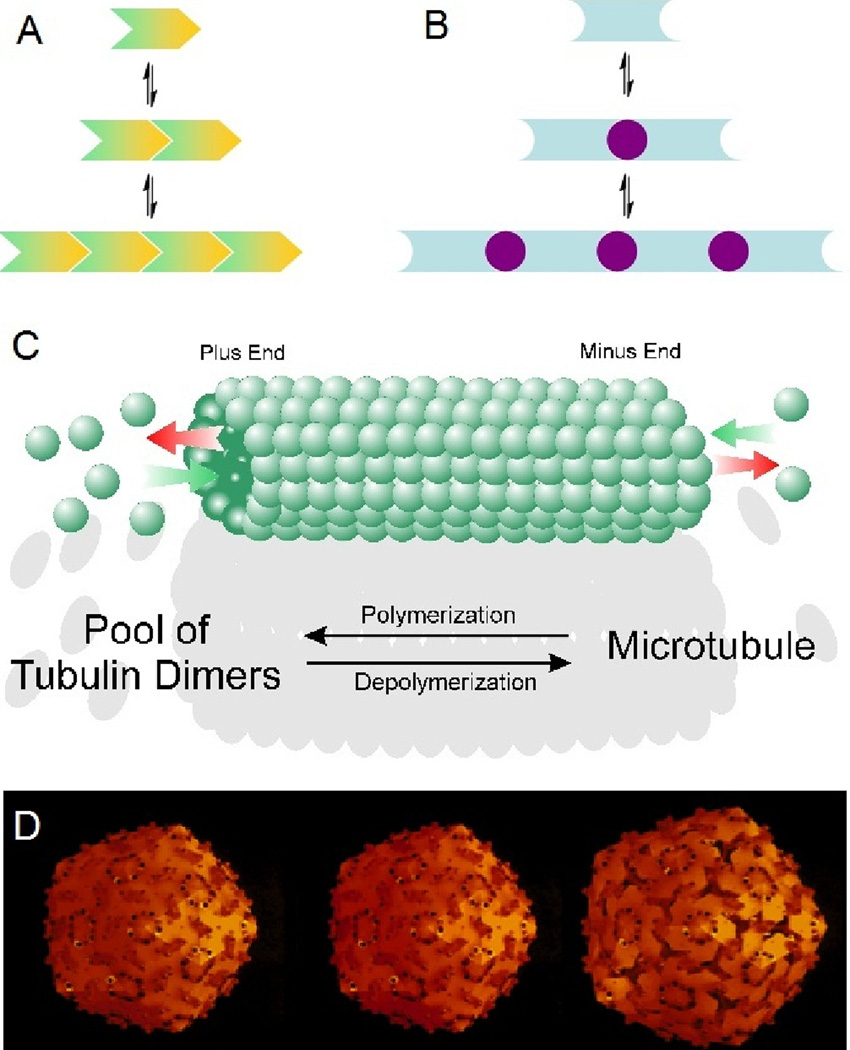
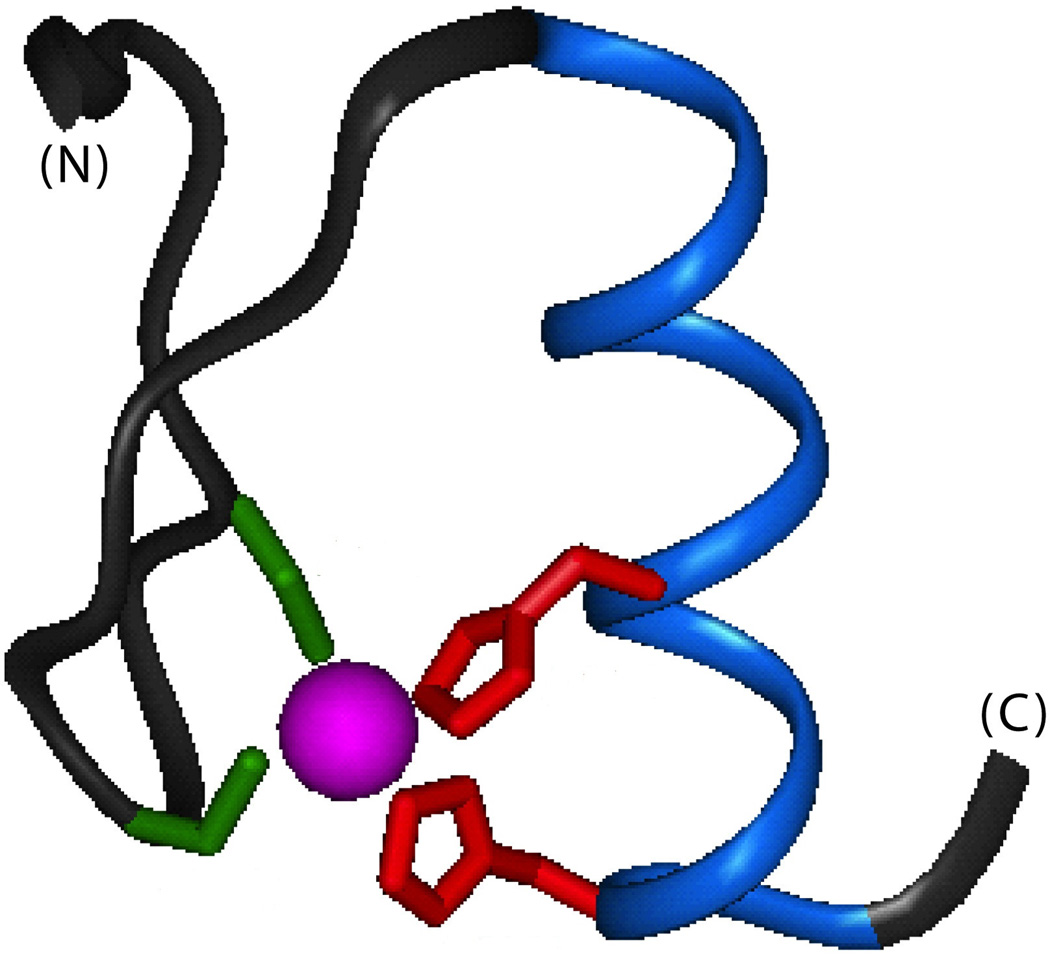
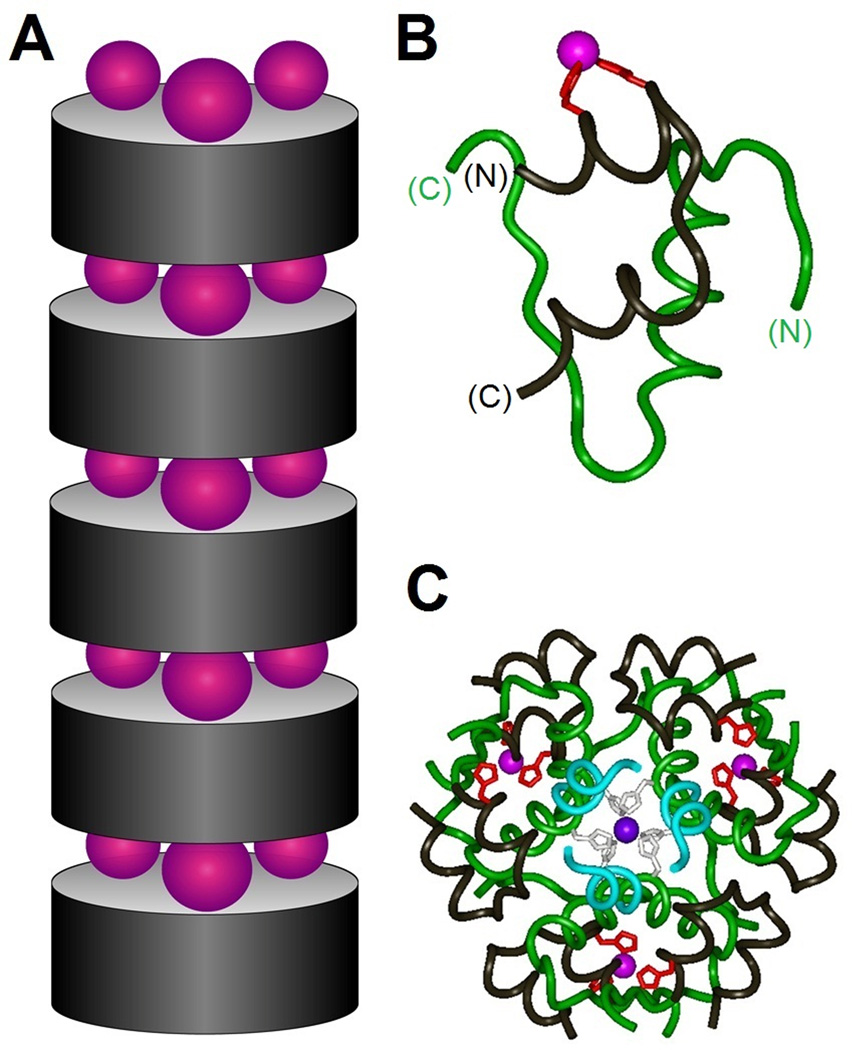
The engineering of insulin analogs represents a triumph of structure-based protein design. A framework has been provided by structures of insulin hexamers. Containing a zinc-coordinated trimer of dimers, such structures represent a storage form of the active insulin monomer. Initial studies focused on destabilization of subunit interfaces. Because disassembly facilitates capillary absorption, such targeted destabilization enabled development of rapid-acting insulin analogs. Converse efforts were undertaken to stabilize the insulin hexamer and promote higher-order self-assembly within the subcutaneous depot toward the goal of enhanced basal glycemic control with reduced risk of hypoglycemia. Current products either operate through isoelectric precipitation (insulin glargine, the active component of Lantus(®); Sanofi-Aventis) or employ an albumin-binding acyl tether (insulin detemir, the active component of Levemir(®); Novo-Nordisk). To further improve pharmacokinetic properties, modified approaches are presently under investigation. Novel strategies have recently been proposed based on subcutaneous supramolecular assembly coupled to (a) large-scale allosteric reorganization of the insulin hexamer (the TR transition), (b) pH-dependent binding of zinc ions to engineered His-X(3)-His sites at hexamer surfaces, or (c) the long-range vision of glucose-responsive polymers for regulated hormone release. Such designs share with wild-type insulin and current insulin products a susceptibility to degradation above room temperature, and so their delivery, storage, and use require the infrastructure of an affluent society. Given the global dimensions of the therapeutic supply chain, we envisage that concurrent engineering of ultra-stable protein analog formulations would benefit underprivileged patients in the developing world.
Conflict of interest statement
The intellectual property pertaining to [HisA4, HisA8]-human insulin and its long-acting formulation is owned by Case Western Reserve University and licensed to Thermalin Diabetes, LLC. M.A.W. holds shares in and is Chief Scientific Officer of Thermalin Diabetes, LLC.; he has also been a consultant to Merck, Inc. and the DEKA Research and Development Corp. A.R.W. is the daughter of M.A.W. D.F.B. is the son of R. Berenson, Chief Executive Office of Thermalin Diabetes, LLC; D.F.B. has an ownership interest in Venzyme Catalyst, LLC, which holds shares in Thermalin Diabetes, LLC. The authors otherwise declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Design of non-standard insulin analogs for the treatment of diabetes mellitus.Curr Diab Rep. 2012 Dec;12(6):697-704. doi: 10.1007/s11892-012-0318-z. Curr Diab Rep. 2012. PMID: 22983891 Free PMC article. Review.
-
Supramolecular protein engineering: design of zinc-stapled insulin hexamers as a long acting depot.J Biol Chem. 2010 Apr 16;285(16):11755-9. doi: 10.1074/jbc.C110.105825. Epub 2010 Feb 24. J Biol Chem. 2010. PMID: 20181952 Free PMC article.
-
Rationale, design, and baseline data of the insulin glargine (Lantus) versus insulin detemir (Levemir) Treat-To-Target (L2T3) study: A multinational, randomized noninferiority trial of basal insulin initiation in type 2 diabetes.Diabetes Technol Ther. 2009 Nov;11(11):739-43. doi: 10.1089/dia.2009.0044. Diabetes Technol Ther. 2009. PMID: 19905891 Clinical Trial.
-
A crossover comparison of glycemic variations in Japanese patients with type 1 diabetes receiving insulin glargine versus insulin detemir twice daily using continuous glucose monitoring (CGM): J COLLECTION (Jikei COmparison of Lantus and LEvemir with Cgm for Thinking Insulin OptimizatioN).Diabetes Technol Ther. 2012 Jul;14(7):596-601. doi: 10.1089/dia.2011.0235. Diabetes Technol Ther. 2012. PMID: 22775404
-
The role of the new basal insulin analogs in addressing unmet clinical needs in people with type 1 and type 2 diabetes.Curr Med Res Opin. 2017 Jun;33(6):1045-1055. doi: 10.1080/03007995.2017.1298522. Epub 2017 Mar 24. Curr Med Res Opin. 2017. PMID: 28277867 Review.
Cited by
-
Display of Single-Chain Insulin-like Peptides on a Yeast Surface.Biochemistry. 2019 Jan 22;58(3):182-188. doi: 10.1021/acs.biochem.8b01094. Epub 2019 Jan 3. Biochemistry. 2019. PMID: 30575376 Free PMC article.
-
Synthesis and Characterization of Phenylboronic Acid-Modified Insulin With Glucose-Dependent Solubility.Front Chem. 2022 Mar 16;10:859133. doi: 10.3389/fchem.2022.859133. eCollection 2022. Front Chem. 2022. PMID: 35372263 Free PMC article.
-
Unraveling Light-Activated Insulin Action in Regulating Blood Glucose: New Photoactivatable Insight as a Novel Modality in Diabetes Management.Molecules. 2024 Mar 14;29(6):1294. doi: 10.3390/molecules29061294. Molecules. 2024. PMID: 38542928 Free PMC article.
-
Insulins for the long term management of diabetes mellitus in dogs: a review.Canine Med Genet. 2022 Feb 14;9(1):1. doi: 10.1186/s40575-022-00114-9. Canine Med Genet. 2022. PMID: 35152907 Free PMC article. Review.
-
Synthesis and Identification of FITC-Insulin Conjugates Produced Using Human Insulin and Insulin Analogues for Biomedical Applications.J Fluoresc. 2016 Mar;26(2):617-29. doi: 10.1007/s10895-015-1748-1. Epub 2015 Dec 12. J Fluoresc. 2016. PMID: 26658795
References
-
- Adams MJ, Blundell TL, Dodson EJ, et al. Structure of rhombohedral 2 zinc insulin crystals. Nature. 1969;224:491–495.
-
- Blundell TL, Cutfield JF, Cutfield SM, et al. Atomic positions in rhombohedral 2-zinc insulin crystals. Nature. 1971;231:506–511. - PubMed
-
- Baker EN, Blundell TL, Cutfield JF, et al. The structure of 2Zn pig insulin crystals at 1.5 Å resolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1988;319:369–456. - PubMed
-
- Brange J, Skelbaek-Pedersen B, Langkjaer L, et al. Galenics of insulin: the physico-chemical and pharmaceutical aspects of insulin and insulin preparations. Berlin: Springer-Verlag; 1987.
-
- Dodson GG, Dodson EJ, Turkenburg JP, et al. Molecular recognition in insulin assembly. Biochem. Soc. Trans. 1993;21:609–614. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
