

Review
doi: 10.1085/jgp.201210795.
Perspectives on: SGP symposium on mitochondrial physiology and medicine: molecular identities of mitochondrial Ca2+ influx mechanism: updated passwords for accessing mitochondrial Ca2+-linked health and disease
Affiliations
- PMID: 22641638
- PMCID: PMC3362516
- DOI: 10.1085/jgp.201210795
Item in Clipboard
Review
Perspectives on: SGP symposium on mitochondrial physiology and medicine: molecular identities of mitochondrial Ca2+ influx mechanism: updated passwords for accessing mitochondrial Ca2+-linked health and disease
J Gen Physiol.
2012 Jun.
No abstract available
Figures
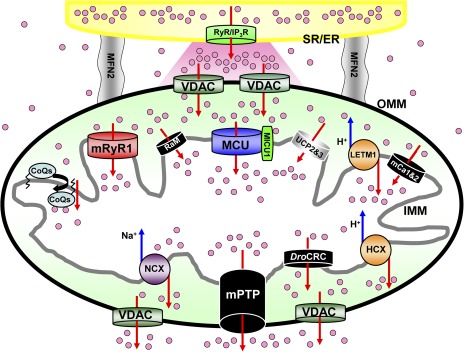
Mitochondrial Ca2+ influx and efflux mechanisms. Schematic diagram of mitochondrial Ca2+ channels/transporters for influx and efflux mechanisms. The functional and morphological tight coupling of ER/SR (yellow) and mitochondria is attributed to the specific structure of inter-organelle tether proteins such as mito-fusion protein 2 (gray) (de Brito and Scorrano, 2008; García-Pérez et al., 2011). Ca2+-releasing sites of ER/SR, IP3 receptors (IP3R), or RyRs (RyR; green) are facing microdomains between mitochondria and ER/SR (shown as a pink region). Ca2+ release from ER/SR dramatically changes [Ca2+]c at this microdomain (Csordás et al., 2010; Giacomello et al., 2010). Then, mitochondria sense the high increases of [Ca2+]c at this microdomain, and [Ca2+]c propagates into the mitochondria matrix through a variety of Ca2+ channels/transporters (Rizzuto et al., 1992). Mitochondrial Ca2+ influx (upper part of this figure) is determined by the MCU (blue) (Baughman et al., 2011; De Stefani et al., 2011), RaM (black) (Sparagna et al., 1995; Buntinas et al., 2001; Bazil and Dash, 2011), HCX (Letm1; orange) (Jiang et al., 2009), mRyR1 (red) (Beutner et al., 2001, 2005), hydroxyl coenzyme Q10 (CoQs) (Bogeski et al., 2011), mCa1 and mCa2 (Michels et al., 2009), and UCP 2 and UPC 3 (Trenker et al., 2007) located at the IMM. MICU1 can bind to Ca2+ by its EF hand, but this protein does not make the channel pore because of its single-transmembrane structure (Perocchi et al., 2010). The mPTP (black) (Giacomello et al., 2007), NCX (purple) (Palty et al., 2010), and HCX (orange) (Jiang et al., 2009) contribute to Ca2+ efflux (lower part of this figure) in mammalian cells. Drosophila mitochondria possess another selective Ca2+ release channel (DroCRC; black) with unique featured characteristics intermediate between the permeability transition pore of yeast and mammals (von Stockum et al., 2011). Letm1 also works as a Ca2+ efflux pathway when [Ca2+]c becomes high (Jiang et al., 2009) (see also Figs. 2 and 3 E). Voltage-dependent anion-selective channels (VDAC; dark green) provide a pathway for Ca2+ and metabolite transport across the outer mitochondrial membrane (OMM). The channels/transporters for which molecular identities are still unknown are shown as black. Red arrows show Ca2+ movements, and blue arrows show other ion movements.
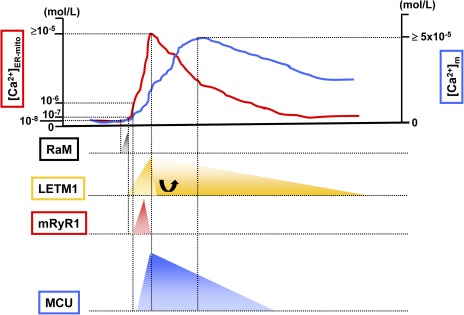
Mitochondrial Ca2+ influx mechanisms during cytosolic/mitochondrial Ca2+ transient. The top of the figure is an example of Ca2+ transient at microdomains between mitochondria and ER/SR ([Ca2+]ER-mito; red line) and Ca2+ transient at the mitochondrial matrix ([Ca2+]m; blue line). RaM (black) shows 50-fold faster Ca2+ transport compared with the MCU, and the activation peak is at 50 nM of extra-mitochondrial Ca2+. Letm1 (orange) can be activated at ≥200 nM of extra-mitochondrial Ca2+, but at high [Ca2+]ER-mito condition, a role of Letm1 shifts to Ca2+ efflux rather than Ca2+ uptake into the mitochondrial matrix. mRyR1 (red) can start to be activated at 1 µM of extra-mitochondrial Ca2+, with a fivefold faster Ca2+ transport compared with the MCU. 2 µM is the half-maximal concentration for Ca2+-dependent activation of mRyR1, and 20 µM is the half-maximal concentration for Ca2+-dependent inhibition. Thus, mRyR1 inactivates before [Ca2+]ER-mito reaches the peak. A lower concentration of extra-mitochondrial Ca2+ (such as 200–300 nM) does not activate MCU, and at least >1 µM Ca2+ is needed for the initial activation. The estimated half-maximal concentration for the activation of MCU is ≅20 mM.
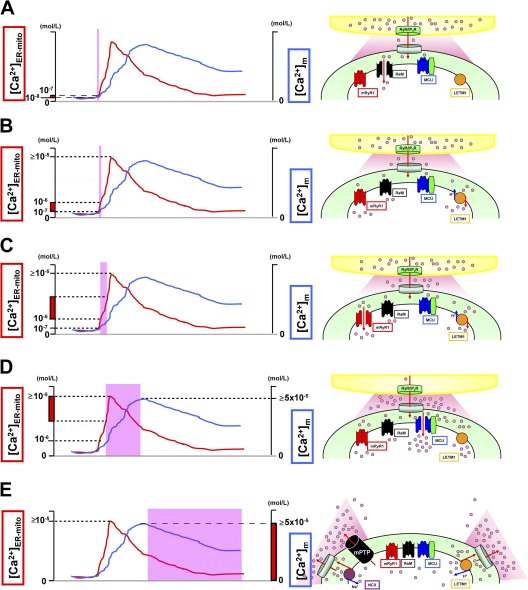
Activation/inactivation patterns of Ca2+ influx/efflux mechanisms. (A) At first, RaM (black) is activated at a very initial phase of [Ca2+]ER-mito transient (<200 nM), with faster Ca2+ uptake kinetics (ms time scale). (B) Letm1 (orange) starts to uptake Ca2+ at ≥200 nM [Ca2+]ER-mito. (C) mRyR1 (red) starts to open at ≅1 µM [Ca2+]ER-mito, with a fivefold faster Ca2+ transport compared with the MCU, and inactivates before [Ca2+]ER-mito reaches the peak. (D) Finally, MCU (blue) starts to activate at >1 µM [Ca2+]ER-mito, and the activity increases in a [Ca2+]ER-mito–dependent manner. At this point, Letm1 (orange) shifts from Ca2+-uptake mode to Ca2+-efflux mode. (E) mPTP (black) and NCX (purple) contribute to Ca2+ efflux in mammalian cells and form the decay phase of [Ca2+]m transient. Letm1 also works as a Ca2+ efflux pathway at this phase. The channels/transporters of which the molecular identities are still unknown are shown as black. Red arrows show Ca2+ movements, and blue arrows show other ion movements.
References
-
- Baughman J.M., Perocchi F., Girgis H.S., Plovanich M., Belcher-Timme C.A., Sancak Y., Bao X.R., Strittmatter L., Goldberger O., Bogorad R.L., et al. 2011. Integrative genomics identifies MCU as an essential component of the mitochondrial calcium uniporter. Nature. 476:341–345 10.1038/nature10234 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
