

Reducing ligation bias of small RNAs in libraries for next generation sequencing
- PMID: 22647250
- PMCID: PMC3489589
- DOI: 10.1186/1758-907X-3-4
Reducing ligation bias of small RNAs in libraries for next generation sequencing
Abstract
Background: The use of nucleic acid-modifying enzymes has driven the rapid advancement in molecular biology. Understanding their function is important for modifying or improving their activity. However, functional analysis usually relies upon low-throughput experiments. Here we present a method for functional analysis of nucleic acid-modifying enzymes using next generation sequencing.
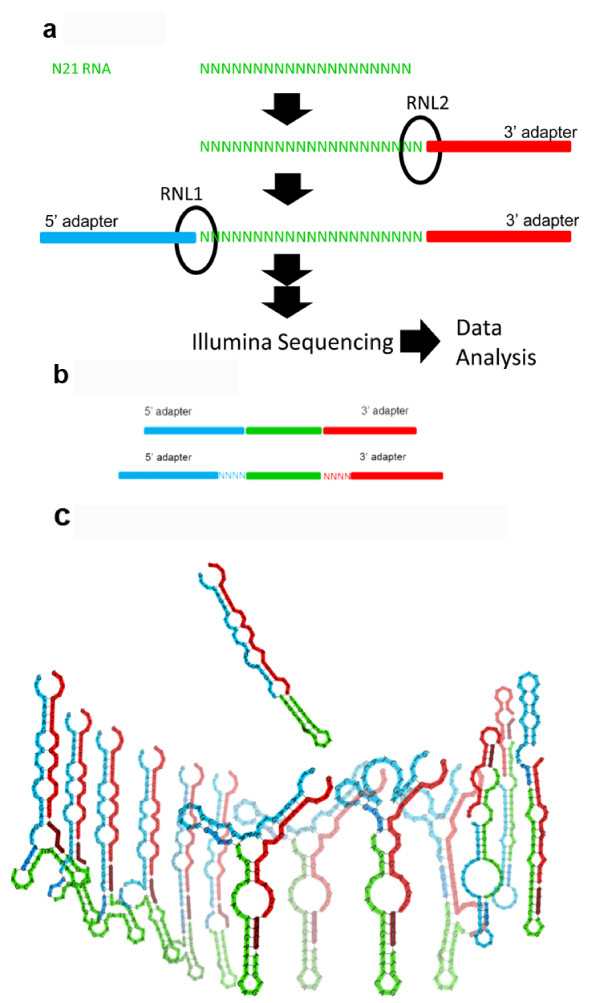
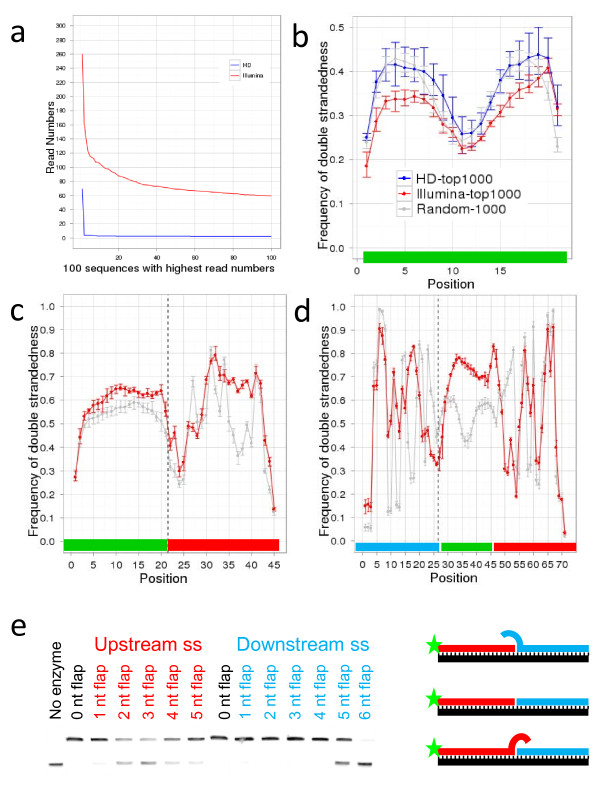
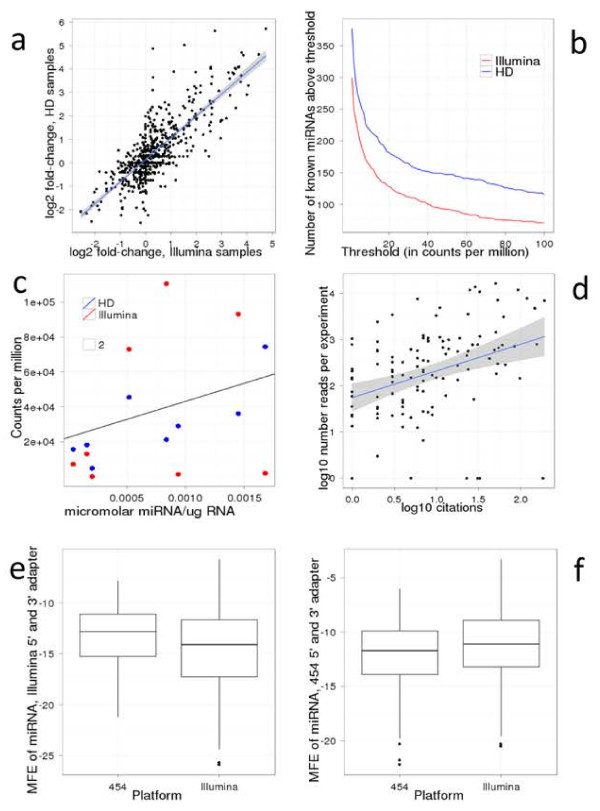
Findings: We demonstrate that sequencing data of libraries generated by RNA ligases can reveal novel secondary structure preferences of these enzymes, which are used in small RNA cloning and library preparation for NGS. Using this knowledge we demonstrate that the cloning bias in small RNA libraries is RNA ligase-dependent. We developed a high definition (HD) protocol that reduces the RNA ligase-dependent cloning bias. The HD protocol doubled read coverage, is quantitative and found previously unidentified microRNAs. In addition, we show that microRNAs in miRBase are those preferred by the adapters of the main sequencing platform.
Conclusions: Sequencing bias of small RNAs partially influenced which microRNAs have been studied in depth; therefore most previous small RNA profiling experiments should be re-evaluated. New microRNAs are likely to be found, which were selected against by existing adapters. Preference of currently used adapters towards known microRNAs suggests that the annotation of all existing small RNAs, including miRNAs, siRNAs and piRNAs, has been biased.
Figures
References
-
- Ho CK, Wang LK, Lima CD, Shuman S. Structure and Mechanism of RNA Ligase. Structure. 2004;12(2):327–339. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases