

Genome-wide patterns of intrahuman dengue virus diversity reveal associations with viral phylogenetic clade and interhost diversity
- PMID: 22647702
- PMCID: PMC3421746
- DOI: 10.1128/JVI.00736-12
Genome-wide patterns of intrahuman dengue virus diversity reveal associations with viral phylogenetic clade and interhost diversity
Abstract
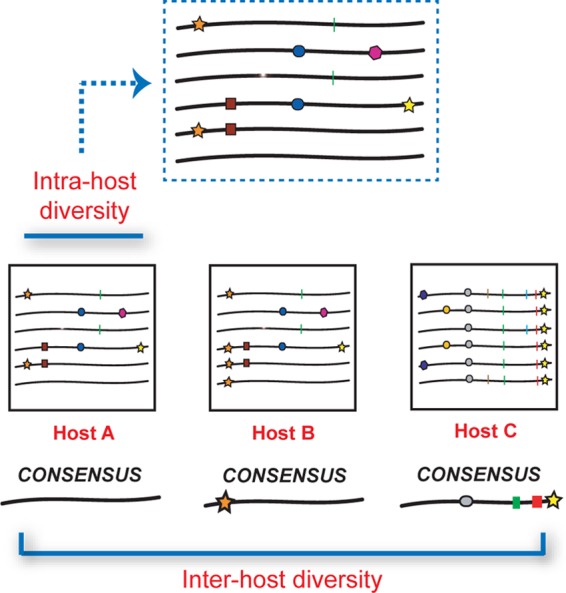
Analogous to observations in RNA viruses such as human immunodeficiency virus, genetic variation associated with intrahost dengue virus (DENV) populations has been postulated to influence viral fitness and disease pathogenesis. Previous attempts to investigate intrahost genetic variation in DENV characterized only a few viral genes or a limited number of full-length genomes. We developed a whole-genome amplification approach coupled with deep sequencing to capture intrahost diversity across the entire coding region of DENV-2. Using this approach, we sequenced DENV-2 genomes from the serum of 22 Nicaraguan individuals with secondary DENV infection and captured ∼75% of the DENV genome in each sample (range, 40 to 98%). We identified and quantified variants using a highly sensitive and specific method and determined that the extent of diversity was considerably lower than previous estimates. Significant differences in intrahost diversity were detected between genes and also between antigenically distinct domains of the Envelope gene. Interestingly, a strong association was discerned between the extent of intrahost diversity in a few genes and viral clade identity. Additionally, the abundance of viral variants within a host, as well as the impact of viral mutations on amino acid encoding and predicted protein function, determined whether intrahost variants were observed at the interhost level in circulating Nicaraguan DENV-2 populations, strongly suggestive of purifying selection across transmission events. Our data illustrate the value of high-coverage genome-wide analysis of intrahost diversity for high-resolution mapping of the relationship between intrahost diversity and clinical, epidemiological, and virological parameters of viral infection.
Figures





References
-
- Aaskov J, Buzacott K, Thu HM, Lowry K, Holmes EC. 2006. Long-term transmission of defective RNA viruses in humans and Aedes mosquitoes. Science 311:236–238 - PubMed
-
- Balmaseda A, et al. 2006. High seroprevalence of antibodies against dengue virus in a prospective study of schoolchildren in Managua, Nicaragua. Trop. Med. Int. Health 11:935–942 - PubMed
-
- Balmaseda A, Sandoval E, Perez L, Gutierrez CM, Harris E. 1999. Application of molecular typing techniques in the 1998 dengue epidemic in Nicaragua. Am. J. Trop. Med. Hyg. 61:893–897 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
