

Modulation by pregnenolone sulfate of filtering properties in the hippocampal trisynaptic circuit
- PMID: 22648992
- PMCID: PMC3434266
- DOI: 10.1002/hipo.22038
Modulation by pregnenolone sulfate of filtering properties in the hippocampal trisynaptic circuit
Abstract
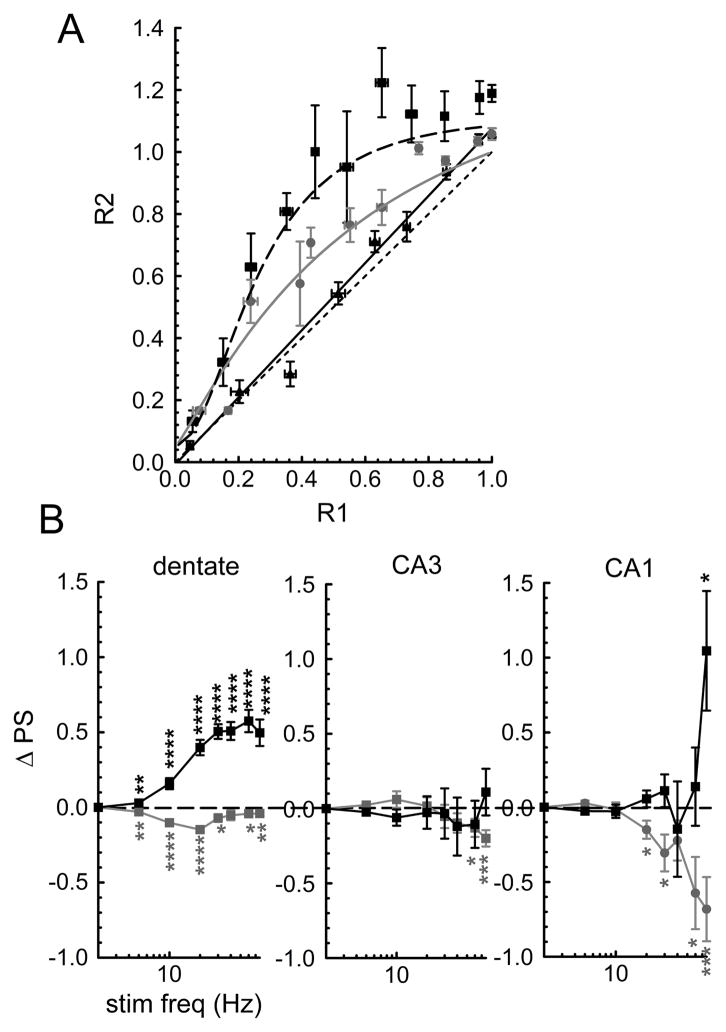
Short-term synaptic plasticity alters synaptic efficacy on a timescale that is relevant to encoding information in spike trains. The dynamics of this plasticity, combined with that of the feedback and feedforward contributions of local interneurons, impose frequency-dependent properties on neuronal networks with implications for nervous system function. The trisynaptic network of the hippocampus is especially well suited to selectively filter components of frequency-dependent signals that are transmitted from the entorhinal cortex. We measured presynaptic [Ca(2+)](i) in perforant path, mossy fiber, or Schaffer collateral terminals while simultaneously measuring field potentials of principal cells of the dentate, CA3, or CA1 synaptic fields over a range of stimulus frequencies of 2 to 77 Hz. In all three synaptic fields, the average [Ca(2+)](i) during a 500 ms stimulus train rose monotonically with stimulus frequency. The average population spike amplitude during this stimulus train, however, exhibited a non-linear relationship to frequency that was distinct for each of the three synaptic fields. The dentate synaptic field exhibited the characteristics of a low pass filter, while both CA synaptic fields had bandpass filter characteristics with a gain that was greater than 1 in the passband frequencies. Importantly, alteration of the dynamic properties of this network could significantly impact information processing performed by the hippocampus. Pregnenolone sulfate (PregS), has frequency-dependent effects on paired- and multipulse plasticity in the dentate and CA1 synaptic fields of the hippocampal formation. We investigated the PregS-dependent modulation of the dynamic properties of transmission by the principal cells of the three hippocampal synaptic fields. Importantly, PregS is capable of altering the pattern separation capabilities that may underlie hippocampal information processing.
Copyright © 2012 Wiley Periodicals, Inc.
Figures






References
-
- Andersen P. Organization of hippocampal neurons and their interconnections. In: Pribram RIK, editor. The Hippocampus Volume I: Structure and Development. New York: Plenum; 1975. pp. 155–175.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
