

Prefrontal neurons represent winning and losing during competitive video shooting games between monkeys
- PMID: 22649245
- PMCID: PMC6703568
- DOI: 10.1523/JNEUROSCI.6479-11.2012
Prefrontal neurons represent winning and losing during competitive video shooting games between monkeys
Abstract
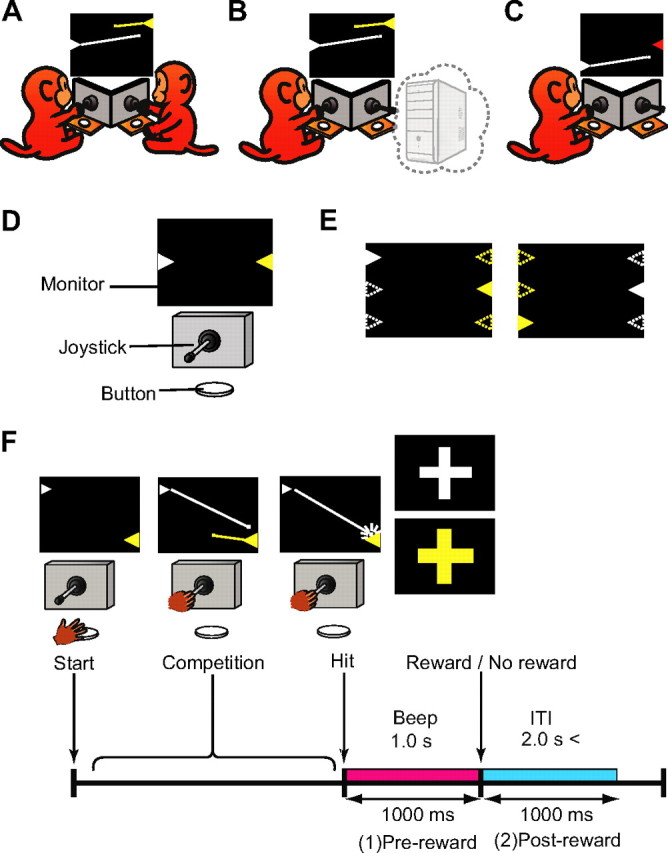
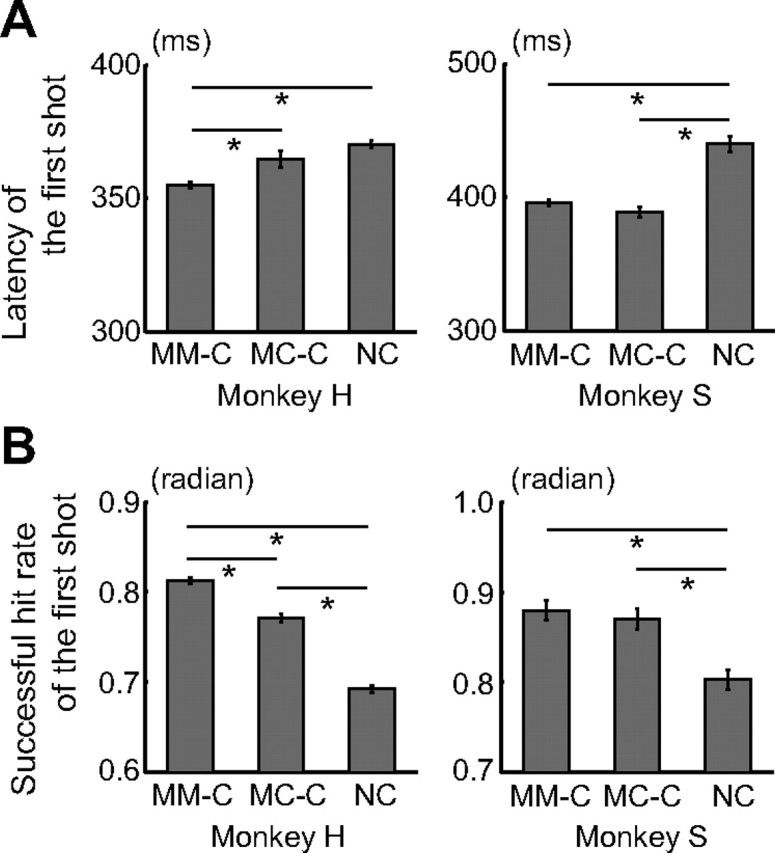
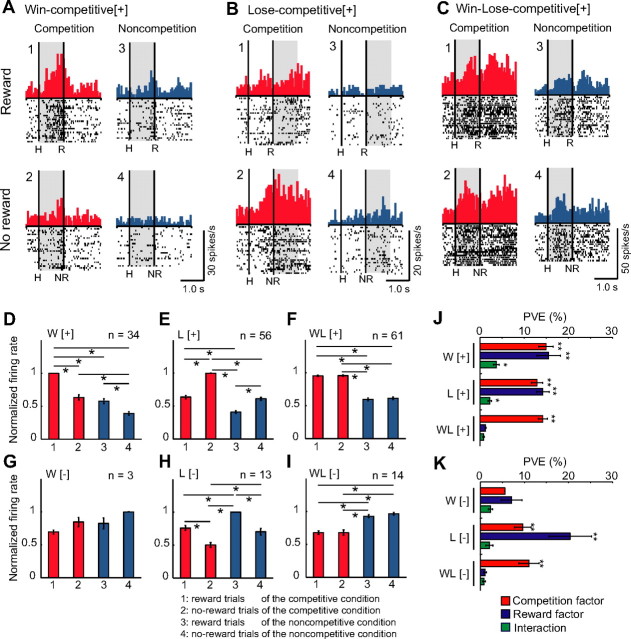
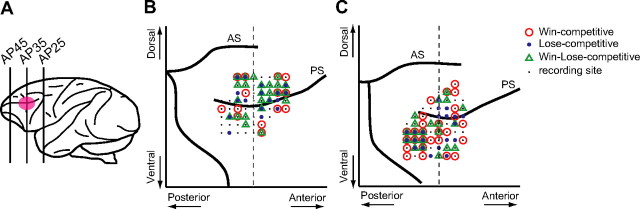
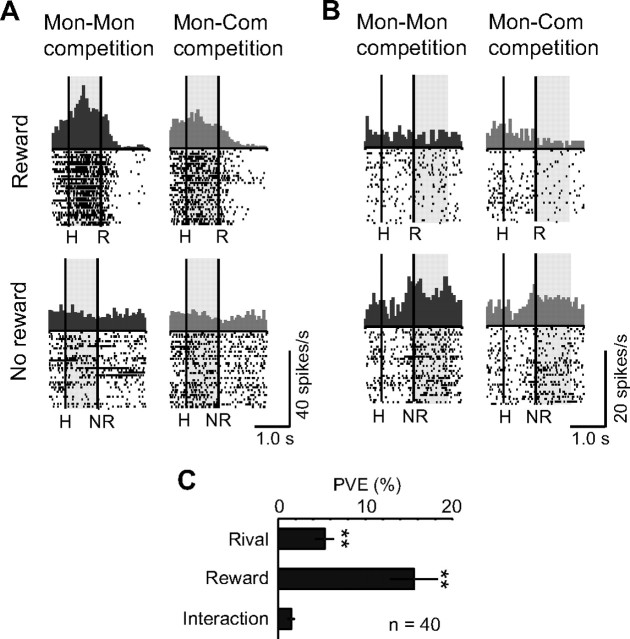
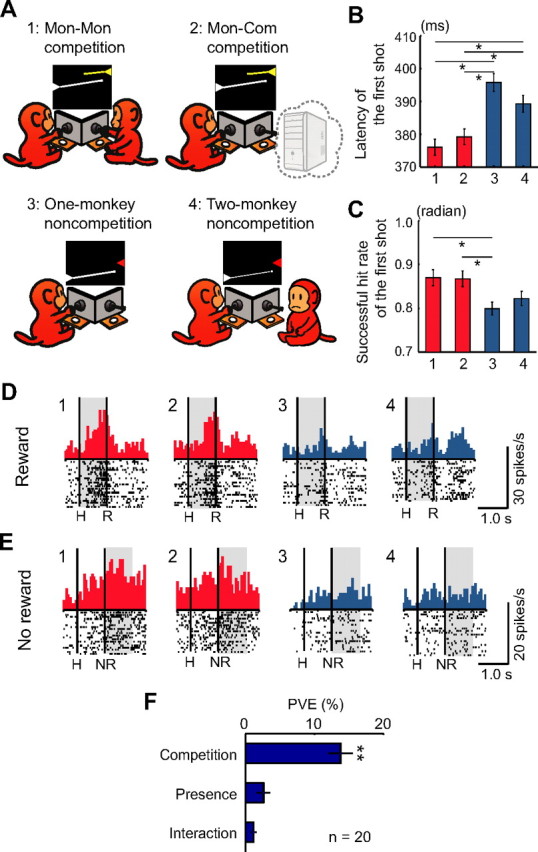
Humans and animals must work to support their survival and reproductive needs. Because resources are limited in the natural environment, competition is inevitable, and competing successfully is vitally important. However, the neuronal mechanisms of competitive behavior are poorly studied. We examined whether neurons in the lateral prefrontal cortex (LPFC) showed response sensitivity related to a competitive game. In this study, monkeys played a video shooting game, either competing with another monkey or the computer, or playing alone without a rival. Monkeys performed more quickly and more accurately in the competitive than in the noncompetitive games, indicating that they were more motivated in the competitive than in the noncompetitive games. LPFC neurons showed differential activity between the competitive and noncompetitive games showing winning- and losing-related activity. Furthermore, activities of prefrontal neurons differed depending on whether the competition was between monkeys or between the monkey and the computer. These results indicate that LPFC neurons may play an important role in monitoring the outcome of competition and enabling animals to adapt their behavior to increase their chances of obtaining a reward in a socially interactive environment.
Figures
References
-
- Amemori K, Sawaguchi T. Rule-dependent shifting of sensorimotor representation in the primate prefrontal cortex. Eur J Neurosci. 2006a;23:1895–1909. - PubMed
-
- Amemori K, Sawaguchi T. Contrasting effects of reward expectation on sensory and motor memories in primate prefrontal neurons. Cereb Cortex. 2006b;16:1002–1015. - PubMed
-
- Barraclough DJ, Conroy ML, Lee D. Prefrontal cortex and decision making in a mixed-strategy game. Nat Neurosci. 2004;7:404–410. - PubMed
-
- Blakemore SJ. The social brain in adolescence. Nat Rev Neurosci. 2008;9:267–277. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources