

Rocket launcher mechanism of collaborative actin assembly defined by single-molecule imaging
- PMID: 22654058
- PMCID: PMC3613992
- DOI: 10.1126/science.1218062
Rocket launcher mechanism of collaborative actin assembly defined by single-molecule imaging
Abstract
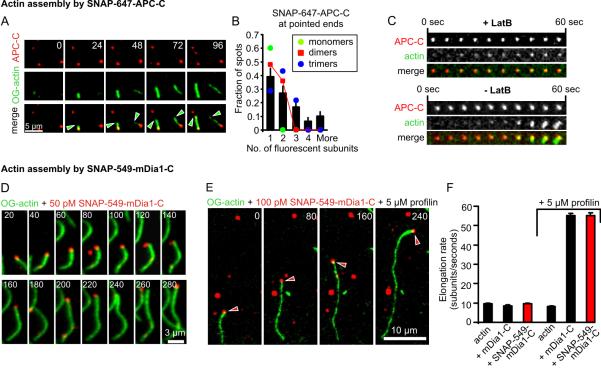
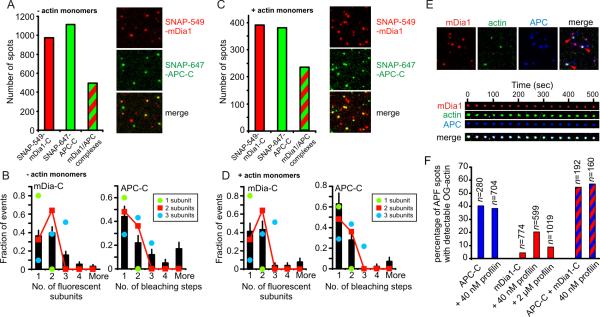
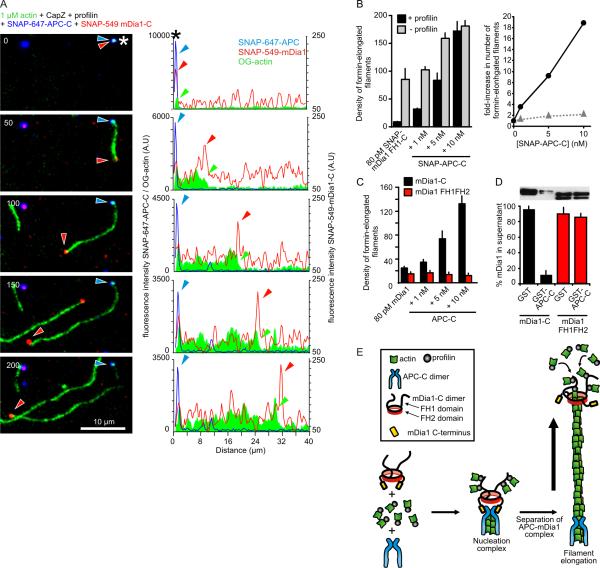
Interacting sets of actin assembly factors work together in cells, but the underlying mechanisms have remained obscure. We used triple-color single-molecule fluorescence microscopy to image the tumor suppressor adenomatous polyposis coli (APC) and the formin mDia1 during filament assembly. Complexes consisting of APC, mDia1, and actin monomers initiated actin filament formation, overcoming inhibition by capping protein and profilin. Upon filament polymerization, the complexes separated, with mDia1 moving processively on growing barbed ends while APC remained at the site of nucleation. Thus, the two assembly factors directly interact to initiate filament assembly and then separate but retain independent associations with either end of the growing filament.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
