

Integrating body and organ size in Drosophila: recent advances and outstanding problems
- PMID: 22654869
- PMCID: PMC3356080
- DOI: 10.3389/fendo.2012.00049
Integrating body and organ size in Drosophila: recent advances and outstanding problems
Abstract
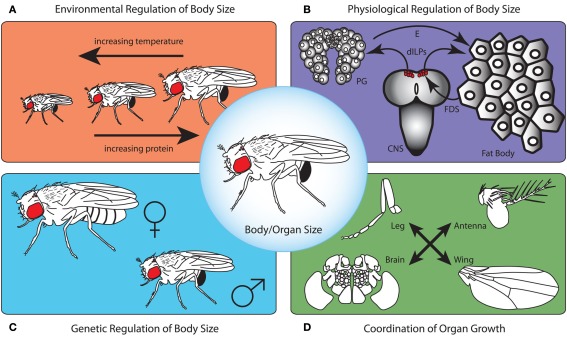
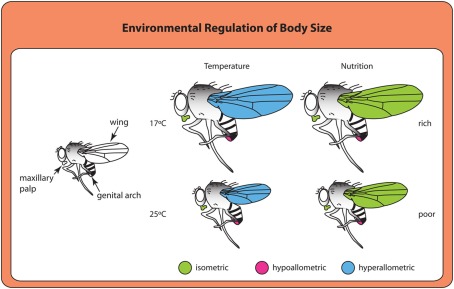
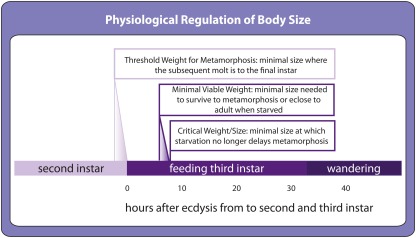
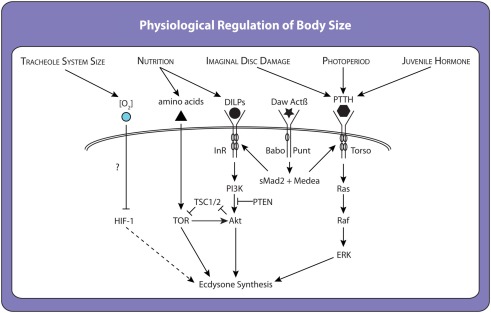
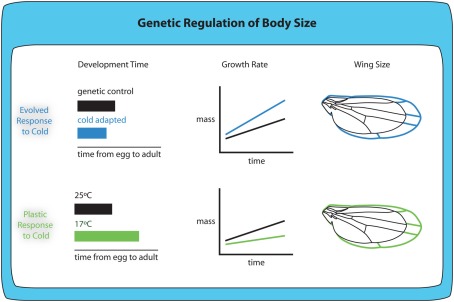
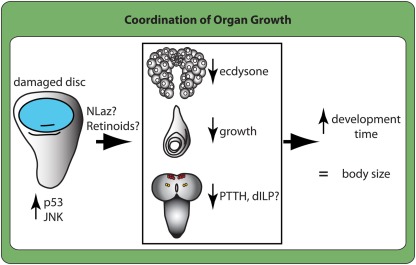
OVER THE PAST TWO DECADES, FUNDAMENTAL STRIDES IN PHYSIOLOGY AND GENETICS HAVE ALLOWED US TO FINALLY GRASP THE DEVELOPMENTAL MECHANISMS REGULATING BODY SIZE, PRIMARILY IN ONE MODEL ORGANISM: the fruit fly Drosophila melanogaster. In Drosophila, as in all animals, final body size is regulated by the rate and duration of growth. These studies have identified important roles for the insulin and the target of rapamycin (TOR) signaling pathways in regulating the growth rate of the larva, the stage most important in determining final adult size. Furthermore, they have shown that the insulin/TOR pathway interacts with hormonal systems, like ecdysone and juvenile hormone, to regulate the timing of development and hence the duration of growth. This interaction allows the growing larvae to integrate cues from the environment with environmentally sensitive developmental windows to ensure that optimal size and proportions are reached given the larval rearing conditions. Results from this work have opened up new avenues of studies, including how environmental cues are integrated to regulate developmental time and how organs maintain proportional growth. Other researchers interested in the evolution of body size are beginning to apply these results to studies of body size evolution and the generation of allometry. With these new findings, and with the developments to come, the field of size control finds itself in the fortunate position of finally being able to tackle century old questions of how organisms achieve final adult size and proportions. This review discusses the state of the art of size control from a Drosophila perspective, and outlines an approach to resolving outstanding issues.
Keywords: developmental timing; ecdysone; environmental effects on body size; genetics of body size and proportions; growth rates; insulin/target of rapamycin signaling; juvenile hormone; regulation of organ growth.
Figures
References
-
- Beadle G. W., Tatum E. L., Clancy C. W. (1938). Food level in relation to rate of development and eye pigmentation in Drosophila melanogaster. Biol. Bull. 75, 447–462 10.2307/1537573 - DOI
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous