

Nanopores: A journey towards DNA sequencing
- PMID: 22658507
- PMCID: PMC3780799
- DOI: 10.1016/j.plrev.2012.05.010
Nanopores: A journey towards DNA sequencing
Abstract
Much more than ever, nucleic acids are recognized as key building blocks in many of life's processes, and the science of studying these molecular wonders at the single-molecule level is thriving. A new method of doing so has been introduced in the mid 1990's. This method is exceedingly simple: a nanoscale pore that spans across an impermeable thin membrane is placed between two chambers that contain an electrolyte, and voltage is applied across the membrane using two electrodes. These conditions lead to a steady stream of ion flow across the pore. Nucleic acid molecules in solution can be driven through the pore, and structural features of the biomolecules are observed as measurable changes in the trans-membrane ion current. In essence, a nanopore is a high-throughput ion microscope and a single-molecule force apparatus. Nanopores are taking center stage as a tool that promises to read a DNA sequence, and this promise has resulted in overwhelming academic, industrial, and national interest. Regardless of the fate of future nanopore applications, in the process of this 16-year-long exploration, many studies have validated the indispensability of nanopores in the toolkit of single-molecule biophysics. This review surveys past and current studies related to nucleic acid biophysics, and will hopefully provoke a discussion of immediate and future prospects for the field.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures
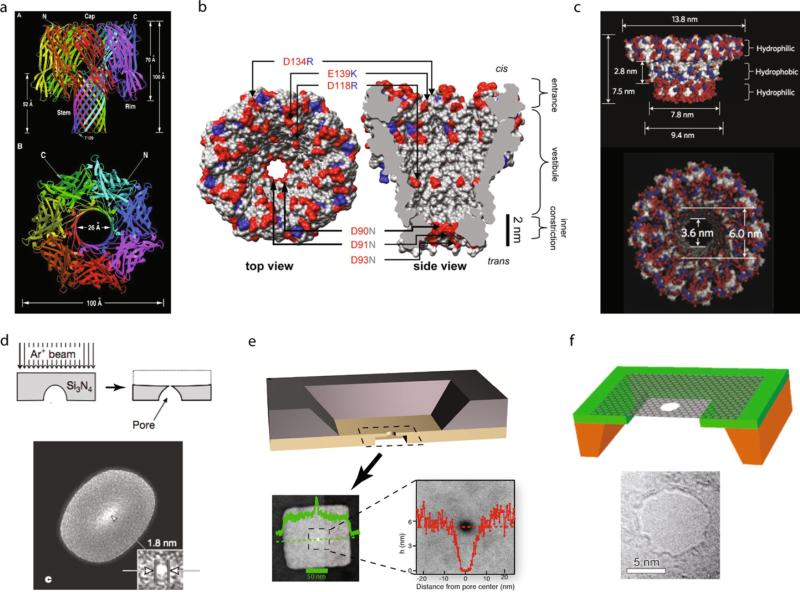
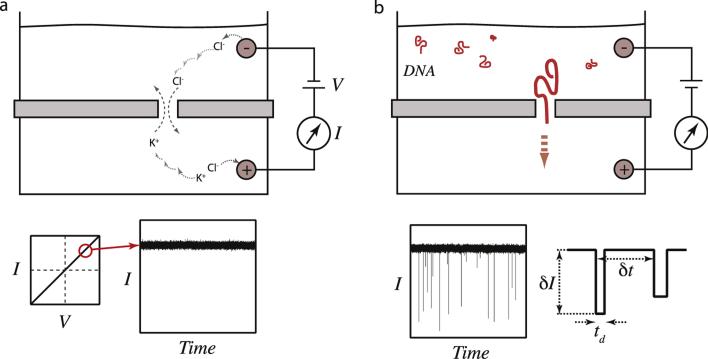
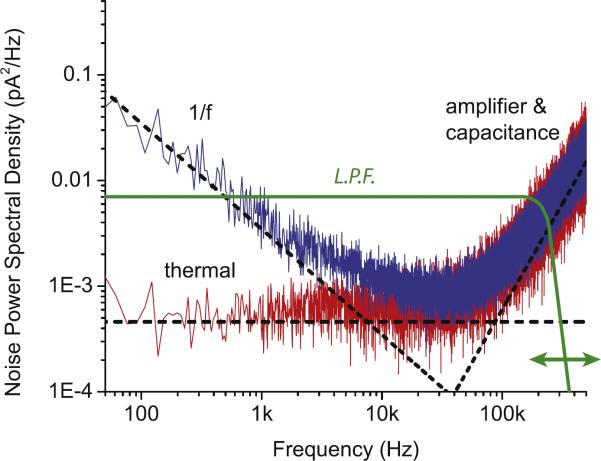



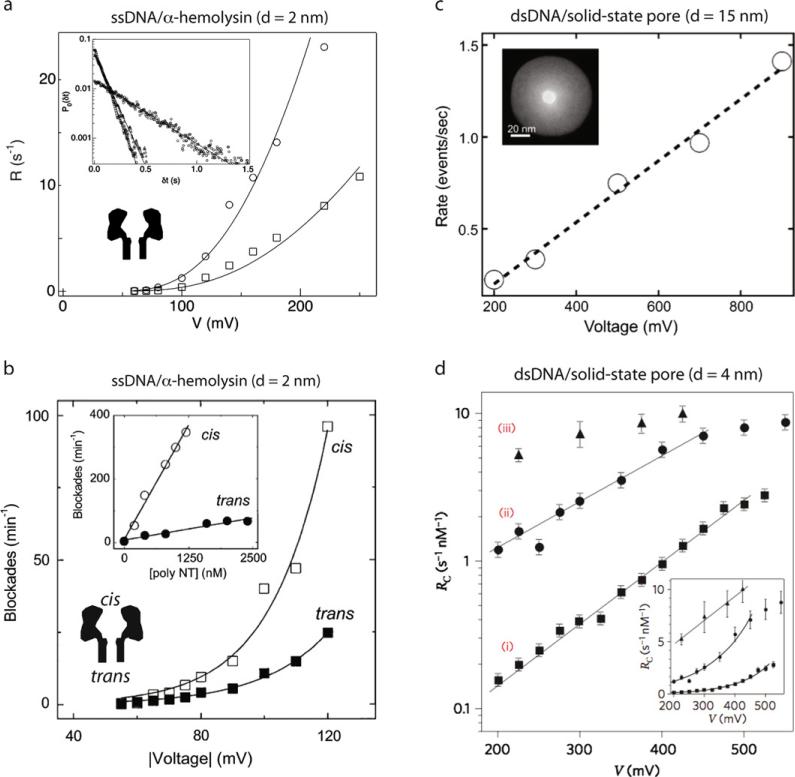
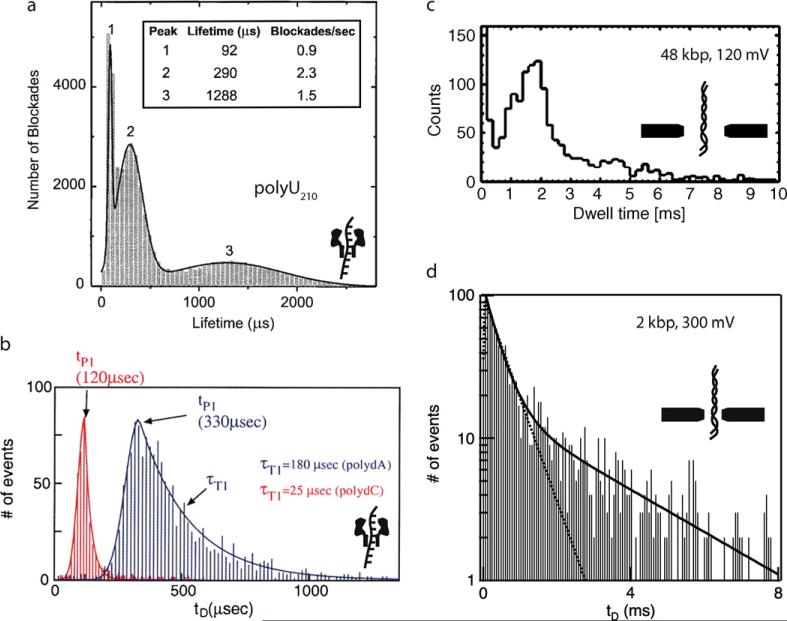


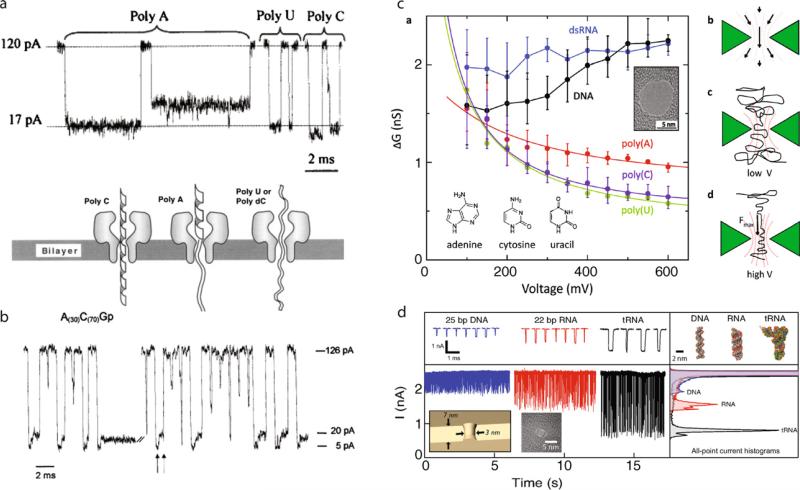


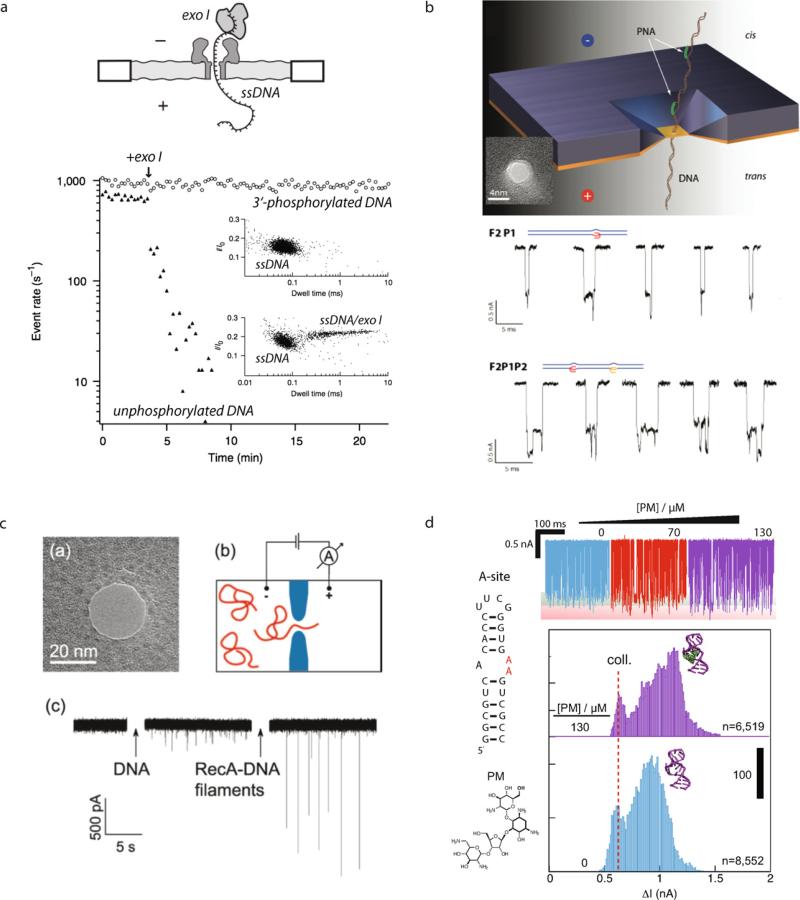



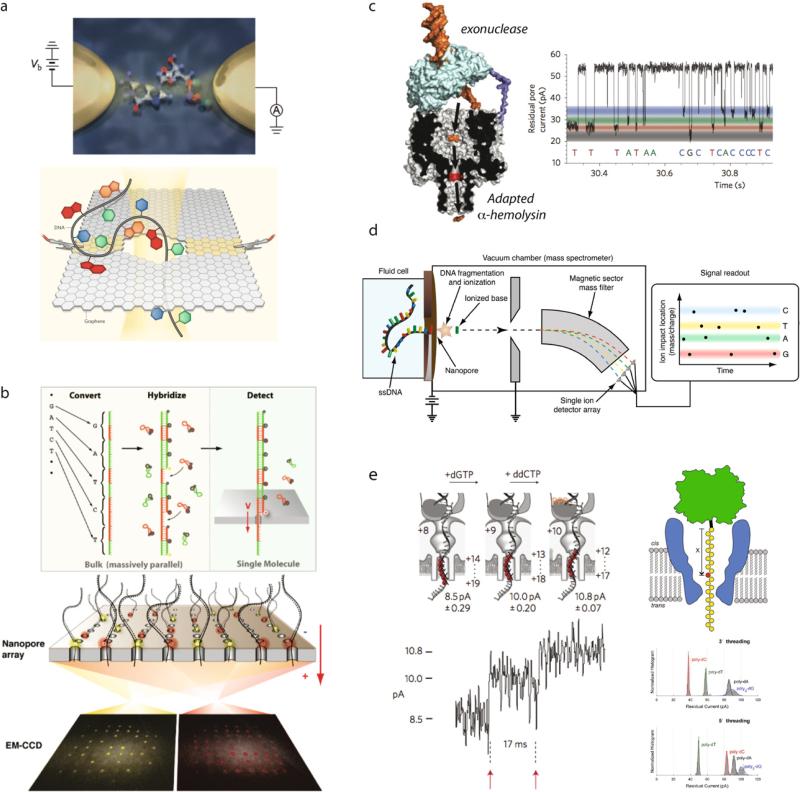
Comment in
-
What about a theory?: Comment on "Nanopores: A journey towards DNA sequencing" by Meni Wanunu.Phys Life Rev. 2012 Jun;9(2):172-3; discussion 174-6. doi: 10.1016/j.plrev.2012.05.011. Epub 2012 May 17. Phys Life Rev. 2012. PMID: 22609434 No abstract available.
-
Bio-inspired nanopore-based sensors: Comment on "Nanopores: A journey towards DNA sequencing" by M. Wanunu.Phys Life Rev. 2012 Jun;9(2):170-1; discussion 174-6. doi: 10.1016/j.plrev.2012.05.012. Epub 2012 May 18. Phys Life Rev. 2012. PMID: 22626932 No abstract available.
-
The richness of the eye of a needle: Comment on "Nanopores: A journey towards DNA sequencing" by M. Wanunu.Phys Life Rev. 2012 Jun;9(2):159-60; discussion 174-6. doi: 10.1016/j.plrev.2012.05.009. Epub 2012 May 17. Phys Life Rev. 2012. PMID: 22626933 No abstract available.
-
DNA nanopore sequencing within the technology landscape: Comment on "Nanopores: A journey towards DNA sequencing" by M. Wanunu.Phys Life Rev. 2012 Jun;9(2):167-9; discussion 174-6. doi: 10.1016/j.plrev.2012.05.013. Epub 2012 May 18. Phys Life Rev. 2012. PMID: 22626935 No abstract available.
-
Nanopores - mission accomplished and what next?: Comment on "Nanopores: A journey towards DNA sequencing" by M. Wanunu.Phys Life Rev. 2012 Jun;9(2):164-6; discussion 174-6. doi: 10.1016/j.plrev.2012.05.014. Epub 2012 May 18. Phys Life Rev. 2012. PMID: 22626937 No abstract available.
-
Are we there yet?: Comment on "Nanopores: A journey towards DNA sequencing" by Meni Wanunu.Phys Life Rev. 2012 Jun;9(2):161-3; discussion 174-6. doi: 10.1016/j.plrev.2012.05.015. Epub 2012 May 22. Phys Life Rev. 2012. PMID: 22652072 No abstract available.
References
-
- Graham MD. The Coulter principle: Foundation of an industry. J Lab Autom. 2003;8:72–81.
-
- Coulter WH. 1953 US Patent #2,656,508.
-
- Deblois RW, Bean CP, Wesley RKA. Electrokinetic measurements with submicron particles and pores by resistive pulse technique. J Colloid Interface Sci. 1977;61:323–35.
-
- Bayley H, Martin CR. Resistive-pulse sensing – From microbes to molecules. Chem Rev. 2000;100:2575–94. - PubMed
-
- Bayley H, Jayasinghe L. Functional engineered channels and pores (Review). Molec Mem Bio. 2004;21:209–20. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
