

Estrogen receptor alpha modulates Toll-like receptor signaling in murine lupus
- PMID: 22659029
- PMCID: PMC3737583
- DOI: 10.1016/j.clim.2012.04.001
Estrogen receptor alpha modulates Toll-like receptor signaling in murine lupus
Abstract
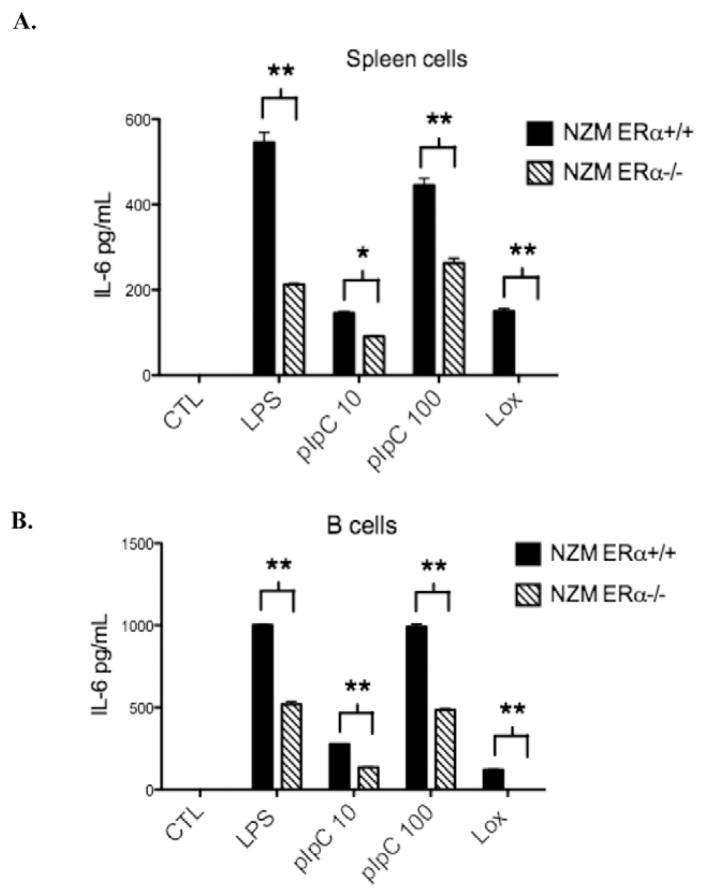
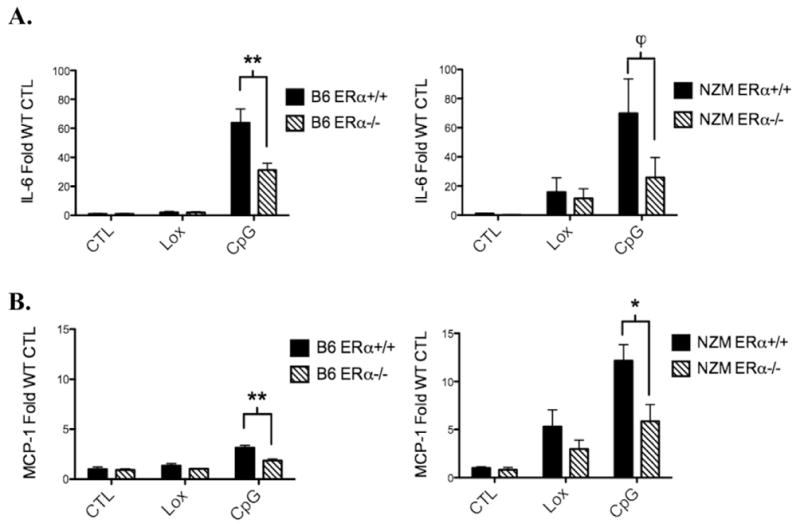
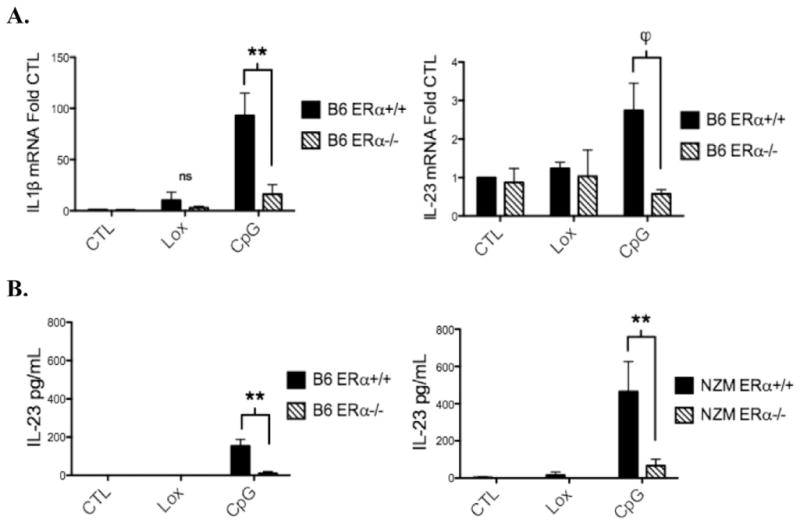
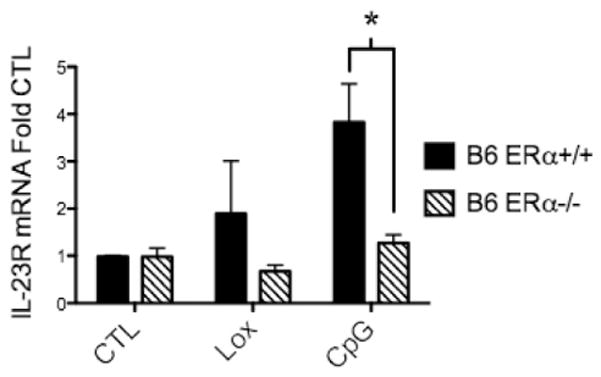
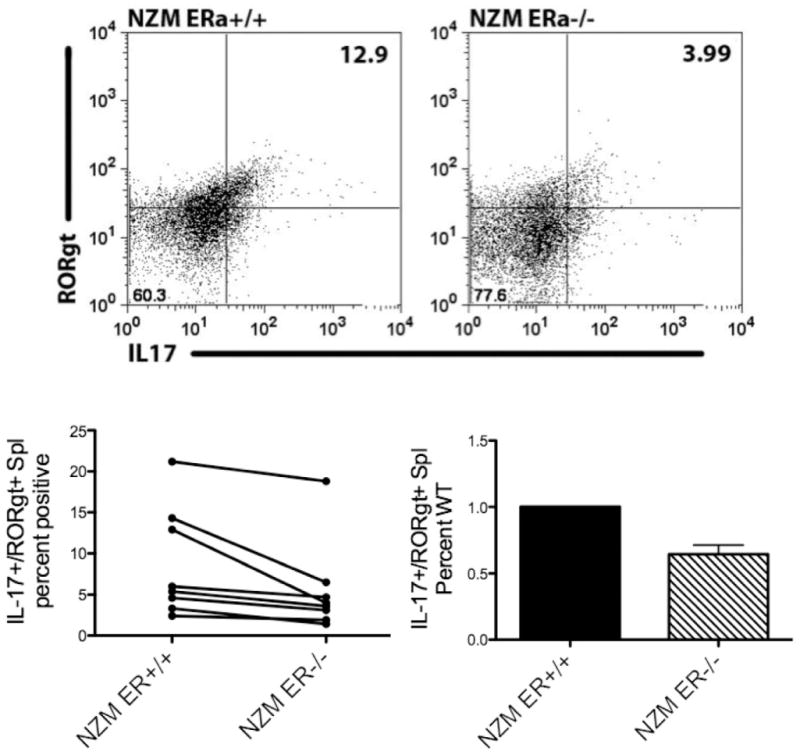
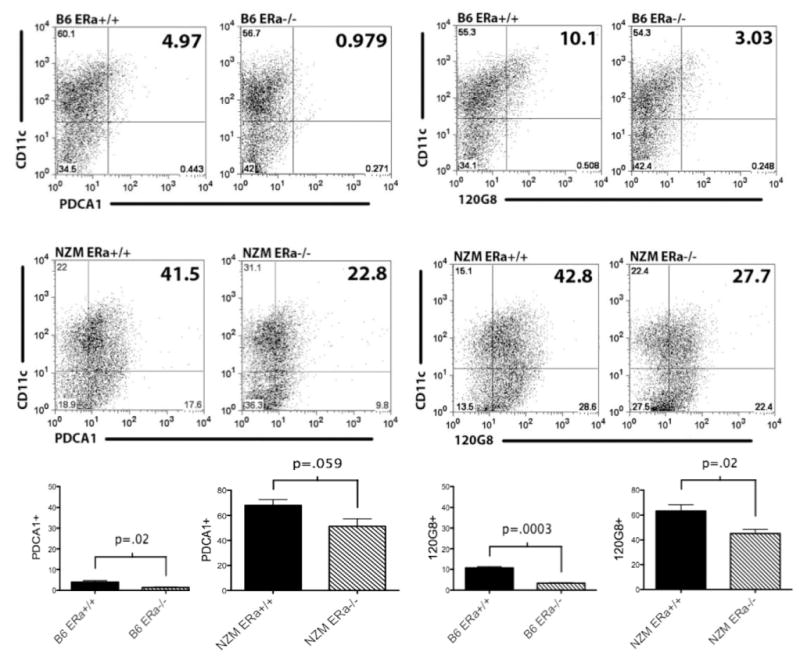
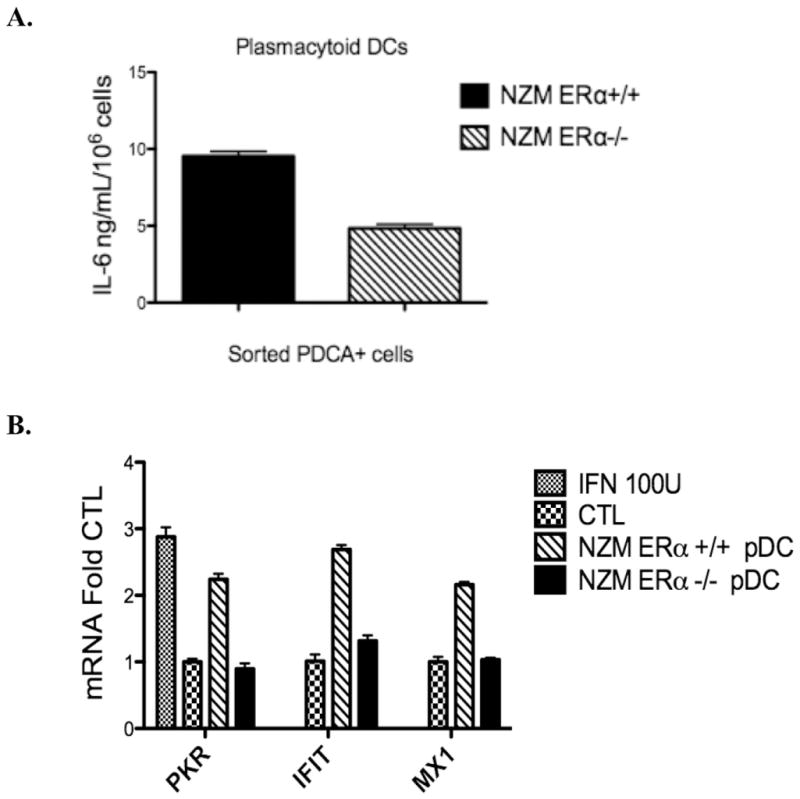
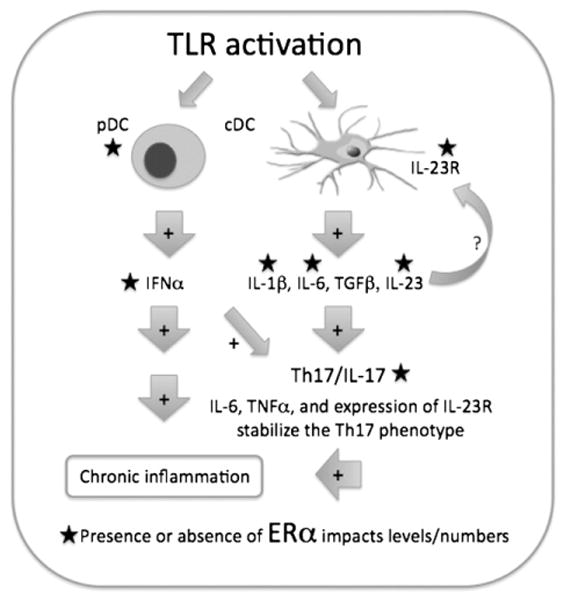
Systemic lupus erythematosus (SLE) is a disease that disproportionately affects females. Despite significant research effort, the mechanisms underlying the female predominance in this disease are largely unknown. Previously, we showed that estrogen receptor alpha knockout (ERαKO) lupus prone female mice had significantly less pathologic renal disease and proteinuria, and significantly prolonged survival. Since autoantibody levels and number and percentage of B/T cells were not significantly impacted by ERα genotype, we hypothesized that the primary benefit of ERα deficiency in lupus nephritis was via modulation of the innate immune response. Using BMDCs and spleen cells/B cells from female wild-type or ERαKO mice, we found that ERαKO-derived cells have a significantly reduced inflammatory response after stimulation with TLR agonists. Our results indicate that the inflammatory response to TLR ligands is significantly impacted by the presence of ERα despite the absence of estradiol, and may partially explain the protective effect of ERα deficiency in lupus-prone animals.
Copyright © 2012. Published by Elsevier Inc.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures
Comment in
-
Why do women get lupus?Clin Immunol. 2012 Jul;144(1):53-6. doi: 10.1016/j.clim.2012.04.003. Epub 2012 Apr 27. Clin Immunol. 2012. PMID: 22659035 No abstract available.
References
-
- Cunningham M, Gilkeson G. Estrogen receptors in immunity and autoimmunity. Clin Rev Allergy Immunol. 2011;40:66–73. - PubMed
-
- Biswas DK, Singh S, Shi Q, Pardee AB, Iglehart JD. Crossroads of estrogen receptor and NF-kappaB signaling. Sci STKE. 2005;2005:pe27. - PubMed
-
- Jakacka M, Ito M, Weiss J, Chien PY, Gehm BD, Jameson JL. Estrogen receptor binding to DNA is not required for its activity through the nonclassical AP1 pathway. J Biol Chem. 2001;276:13615–13621. - PubMed
-
- Safe S. Transcriptional activation of genes by 17 beta-estradiol through estrogen receptor-Sp1 interactions. Vitam Horm. 2001;62:231–252. - PubMed
-
- Lambert KC, Curran EM, Judy BM, Lubahn DB, Estes DM. Estrogen receptor-alpha deficiency promotes increased TNF-alpha secretion and bacterial killing by murine macrophages in response to microbial stimuli in vitro. J Leukoc Biol. 2004;75:1166–1172. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
