

Alternating access mechanism in the POT family of oligopeptide transporters
- PMID: 22659829
- PMCID: PMC3419923
- DOI: 10.1038/emboj.2012.157
Alternating access mechanism in the POT family of oligopeptide transporters
Abstract
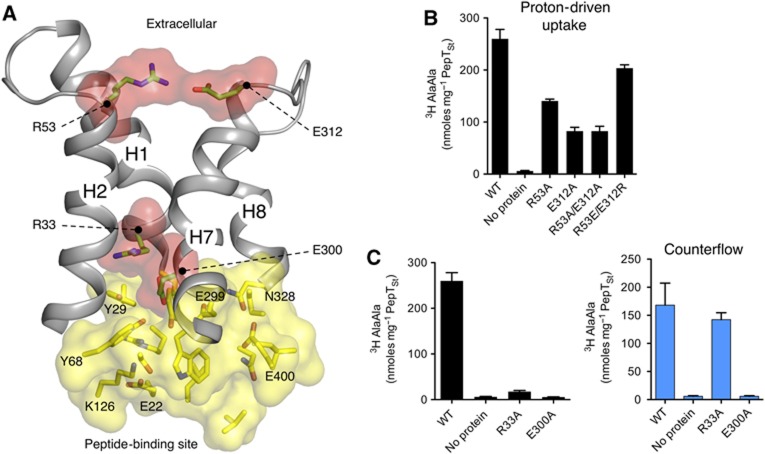
Short chain peptides are actively transported across membranes as an efficient route for dietary protein absorption and for maintaining cellular homeostasis. In mammals, peptide transport occurs via PepT1 and PepT2, which belong to the proton-dependent oligopeptide transporter, or POT family. The recent crystal structure of a bacterial POT transporter confirmed that they belong to the major facilitator superfamily of secondary active transporters. Despite the functional characterization of POT family members in bacteria, fungi and mammals, a detailed model for peptide recognition and transport remains unavailable. In this study, we report the 3.3-Å resolution crystal structure and functional characterization of a POT family transporter from the bacterium Streptococcus thermophilus. Crystallized in an inward open conformation the structure identifies a hinge-like movement within the C-terminal half of the transporter that facilitates opening of an intracellular gate controlling access to a central peptide-binding site. Our associated functional data support a model for peptide transport that highlights the importance of salt bridge interactions in orchestrating alternating access within the POT family.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures




Comment in
-
A POTluck of peptide transporters.EMBO J. 2012 Aug 15;31(16):3382-3. doi: 10.1038/emboj.2012.206. Epub 2012 Jul 27. EMBO J. 2012. PMID: 22842786 Free PMC article.
References
-
- Abrahams JP, Leslie AGW (1996) Methods used in the structure determination of bovine mitochondrial F1 ATPase. Acta Crystallogr D Biol Crystallogr 52: 30–42 - PubMed
-
- Abramson J, Smirnova I, Kasho V, Verner G, Kaback HR, Iwata S (2003) Structure and mechanism of the lactose permease of Escherichia coli. Science 301: 610–615 - PubMed
-
- Anderson CMH, Thwaites DT (2010) Hijacking solute carriers for proton-coupled drug transport. Physiology 25: 364–377 - PubMed
-
- Biegel A, Knutter I, Hartrodt B, Gebauer S, Theis S, Luckner P, Kottra G, Rastetter M, Zebisch K, Thondorf I, Daniel H, Neubert K, Brandsch M (2006) The renal type H+/peptide symporter PEPT2: structure-affinity relationships. Amino Acids 31: 137–156 - PubMed
-
- Blanc E, Roversi P, Vonrhein C, Flensburg C, Lea SM, Bricogne G (2004) Refinement of severely incomplete structures with maximum likelihood in BUSTER-TNT. Acta Crystallogr D Biol Crystallogr 60: 2210–2221 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
