

Neutrophil cerebrovascular transmigration triggers rapid neurotoxicity through release of proteases associated with decondensed DNA
- PMID: 22661091
- PMCID: PMC3381844
- DOI: 10.4049/jimmunol.1200409
Neutrophil cerebrovascular transmigration triggers rapid neurotoxicity through release of proteases associated with decondensed DNA
Abstract
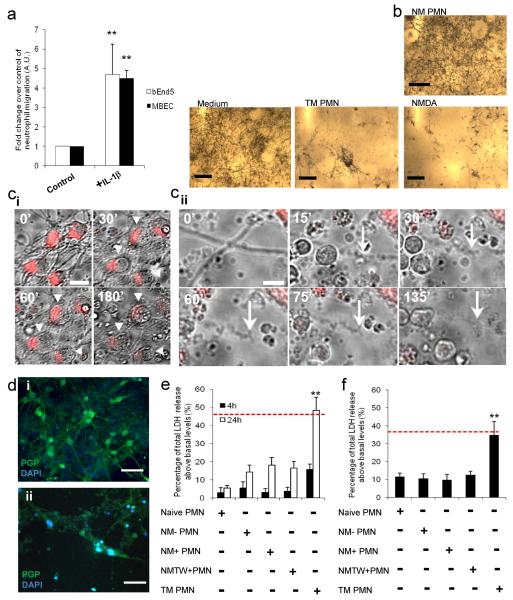
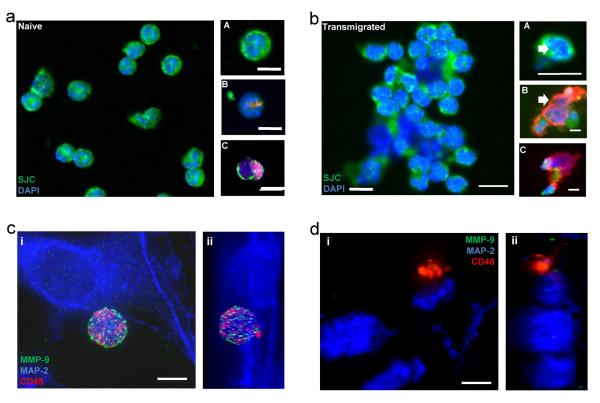
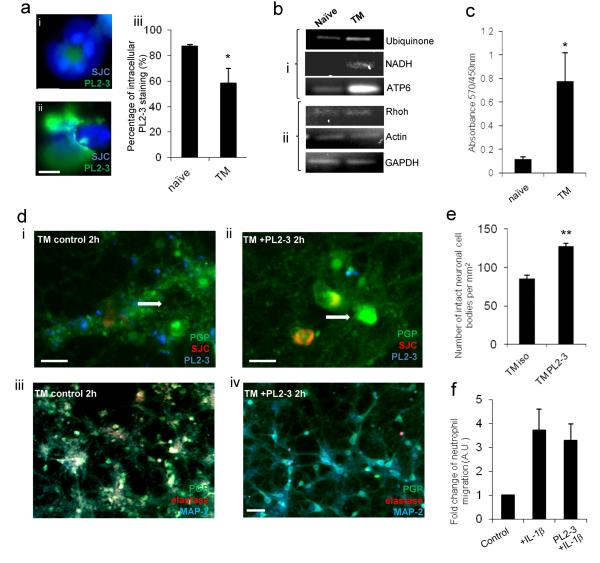
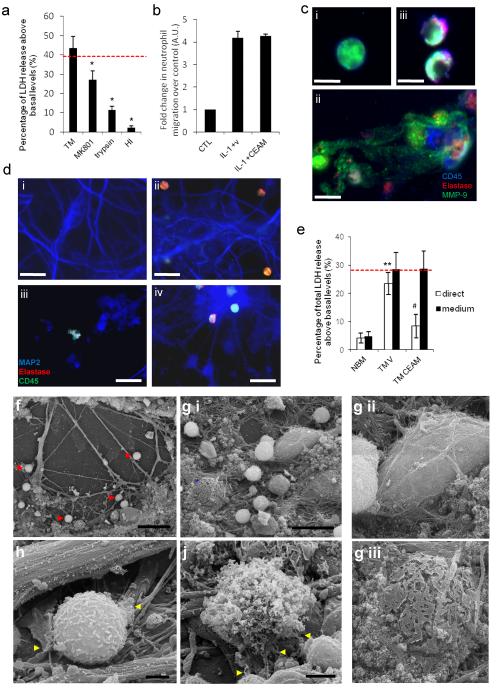
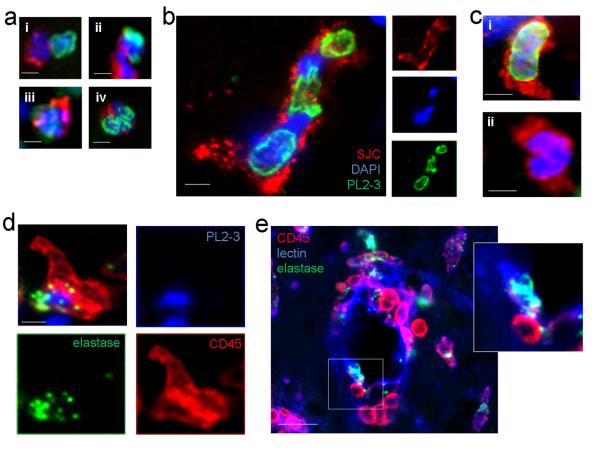
Cerebrovascular inflammation contributes to diverse CNS disorders through mechanisms that are incompletely understood. The recruitment of neutrophils to the brain can contribute to neurotoxicity, particularly during acute brain injuries, such as cerebral ischemia, trauma, and seizures. However, the regulatory and effector mechanisms that underlie neutrophil-mediated neurotoxicity are poorly understood. In this study, we show that mouse neutrophils are not inherently toxic to neurons but that transendothelial migration across IL-1-stimulated brain endothelium triggers neutrophils to acquire a neurotoxic phenotype that causes the rapid death of cultured neurons. Neurotoxicity was induced by the addition of transmigrated neutrophils or conditioned medium, taken from transmigrated neutrophils, to neurons and was partially mediated by excitotoxic mechanisms and soluble proteins. Transmigrated neutrophils also released decondensed DNA associated with proteases, which are known as neutrophil extracellular traps. The blockade of histone-DNA complexes attenuated transmigrated neutrophil-induced neuronal death, whereas the inhibition of key neutrophil proteases in the presence of transmigrated neutrophils rescued neuronal viability. We also show that neutrophil recruitment in the brain is IL-1 dependent, and release of proteases and decondensed DNA from recruited neutrophils in the brain occurs in several in vivo experimental models of neuroinflammation. These data reveal new regulatory and effector mechanisms of neutrophil-mediated neurotoxicity (i.e., the release of proteases and decondensed DNA triggered by phenotypic transformation during cerebrovascular transmigration). Such mechanisms have important implications for neuroinflammatory disorders, notably in the development of antileukocyte therapies.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
