

SUMO-targeted ubiquitin E3 ligase RNF4 is required for the response of human cells to DNA damage
- PMID: 22661230
- PMCID: PMC3371408
- DOI: 10.1101/gad.189274.112
SUMO-targeted ubiquitin E3 ligase RNF4 is required for the response of human cells to DNA damage
Abstract
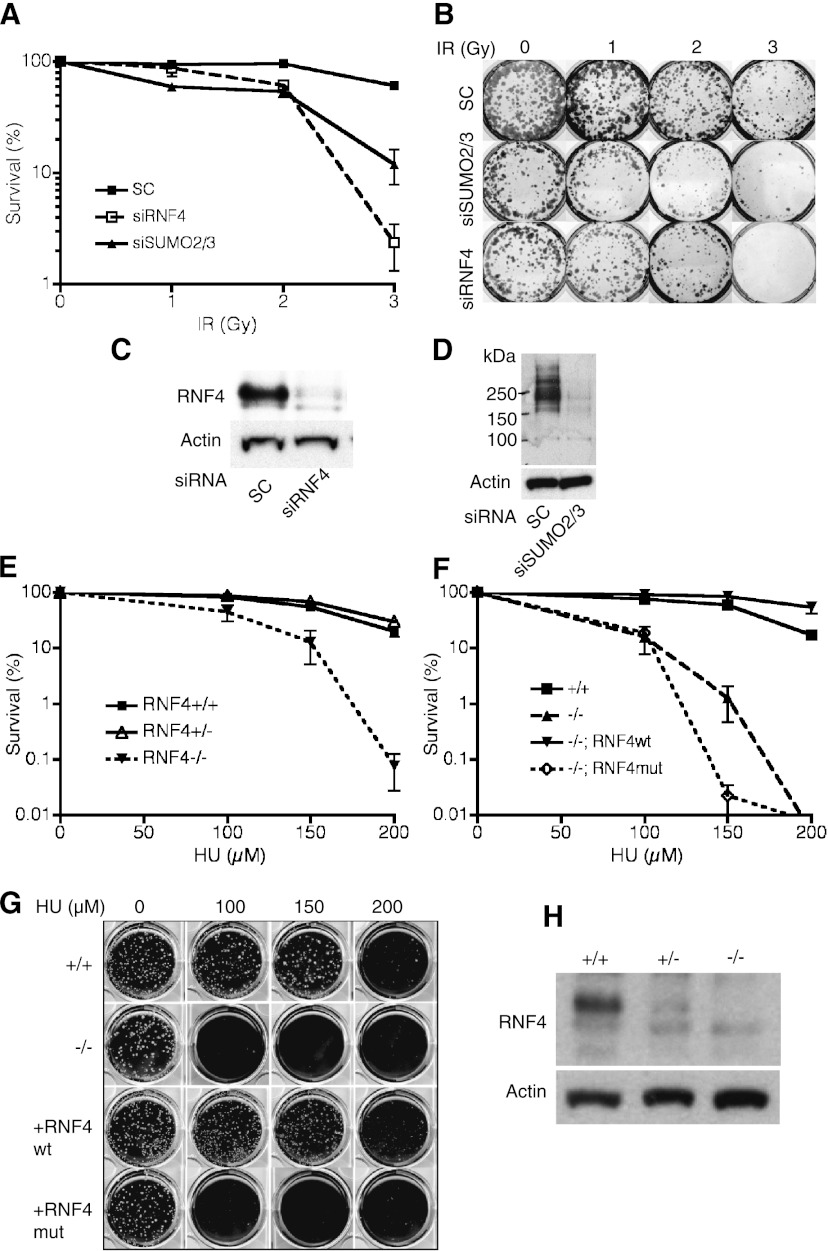
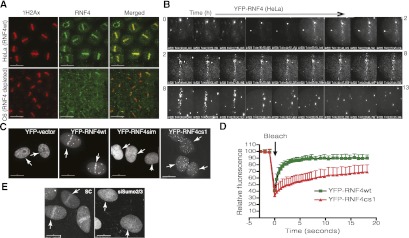
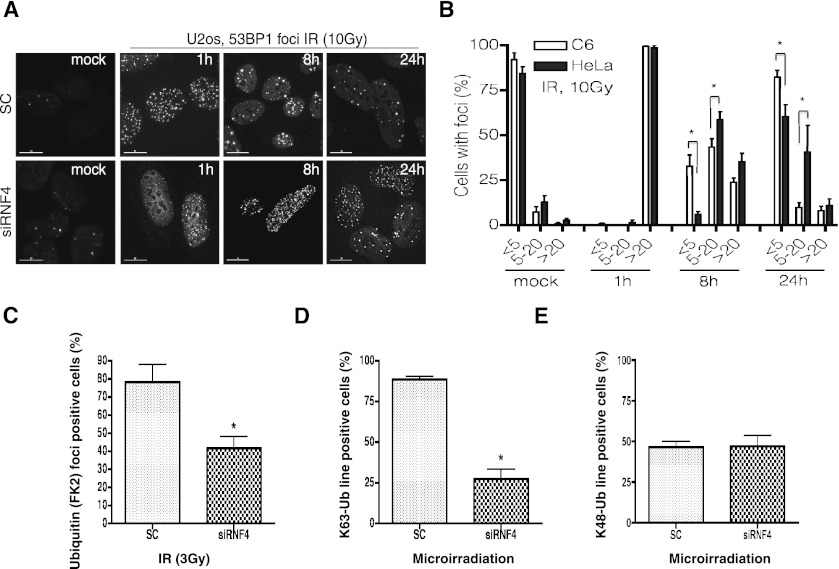
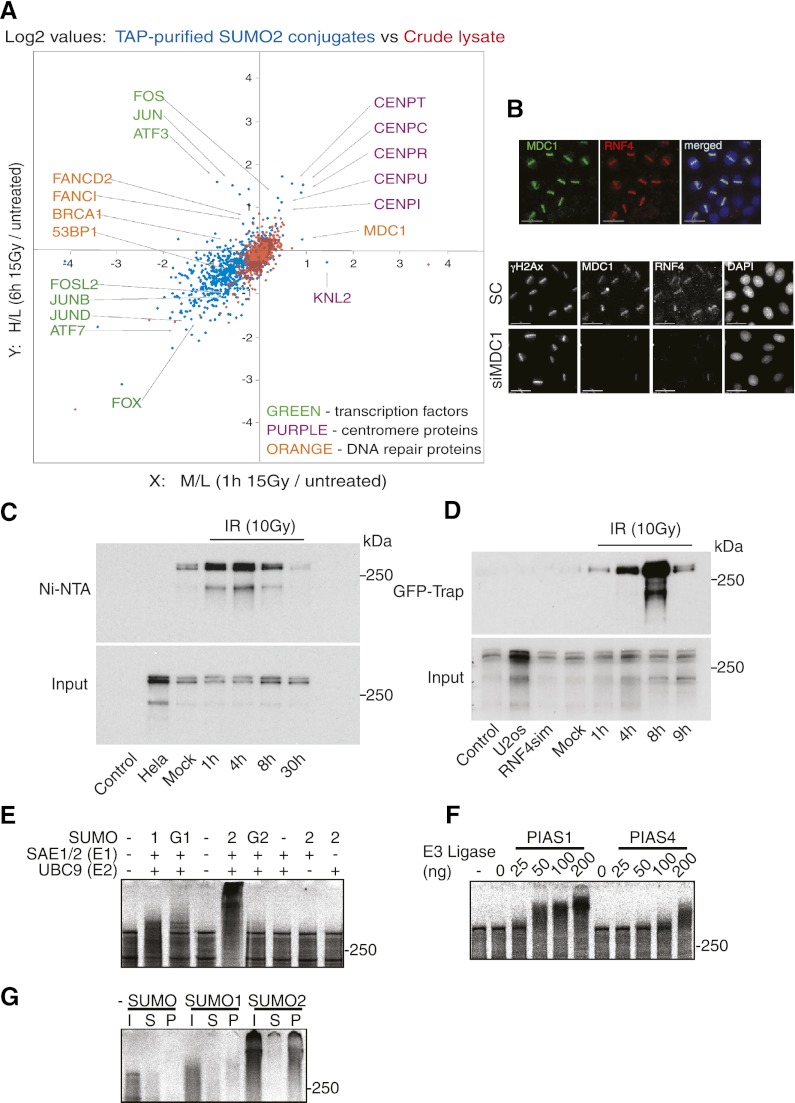
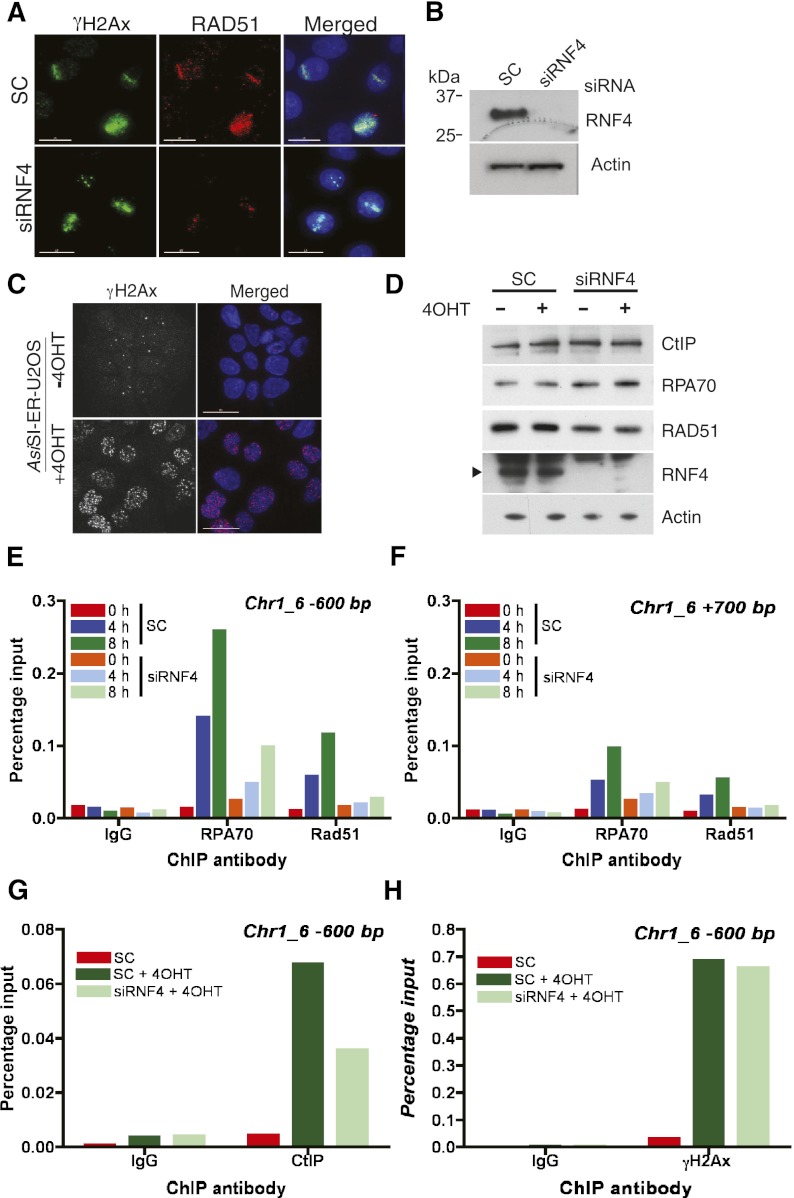
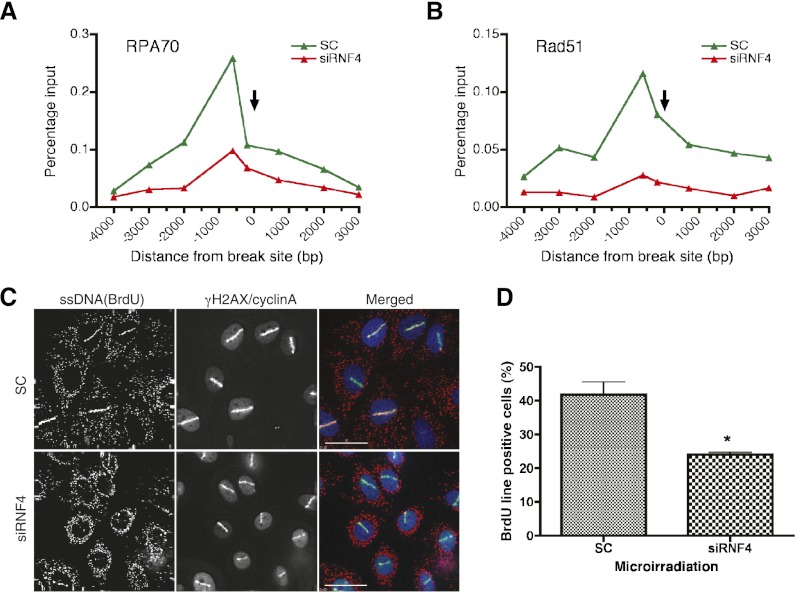
Here we demonstrate that RNF4, a highly conserved small ubiquitin-like modifier (SUMO)-targeted ubiquitin E3 ligase, plays a critical role in the response of mammalian cells to DNA damage. Human cells in which RNF4 expression was ablated by siRNA or chicken DT40 cells with a homozygous deletion of the RNF4 gene displayed increased sensitivity to DNA-damaging agents. Recruitment of RNF4 to double-strand breaks required its RING and SUMO interaction motif (SIM) domains and DNA damage factors such as NBS1, mediator of DNA damage checkpoint 1 (MDC1), RNF8, 53BP1, and BRCA1. In the absence of RNF4, these factors were still recruited to sites of DNA damage, but 53BP1, RNF8, and RNF168 displayed delayed clearance from such foci. SILAC-based proteomics of SUMO substrates revealed that MDC1 was SUMO-modified in response to ionizing radiation. As a consequence of SUMO modification, MDC1 recruited RNF4, which mediated ubiquitylation at the DNA damage site. Failure to recruit RNF4 resulted in defective loading of replication protein A (RPA) and Rad51 onto ssDNA. This appeared to be a consequence of reduced recruitment of the CtIP nuclease, resulting in inefficient end resection. Thus, RNF4 is a novel DNA damage-responsive protein that plays a role in homologous recombination and integrates SUMO modification and ubiquitin signaling in the cellular response to genotoxic stress.
Figures
References
-
- Bergink S, Jentsch S 2009. Principles of ubiquitin and SUMO modifications in DNA repair. Nature 458: 461–467 - PubMed
-
- Boulton SJ 2006. Cellular functions of the BRCA tumour-suppressor proteins. Biochem Soc Trans 34: 633–643 - PubMed
-
- Doil C, Mailand N, Bekker-Jensen S, Menard P, Larsen DH, Pepperkok R, Ellenberg J, Panier S, Durocher D, Bartek J, et al. 2009. RNF168 binds and amplifies ubiquitin conjugates on damaged chromosomes to allow accumulation of repair proteins. Cell 136: 435–446 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous